பின் கிளினிக் ஆக்ஸிஜனேற்ற அழுத்த உடலியக்க மற்றும் செயல்பாட்டு மருத்துவக் குழு. ஆக்ஸிஜனேற்ற அழுத்தம் என்பது எதிர்வினை ஆக்ஸிஜன் (ஃப்ரீ ரேடிக்கல்கள்) மற்றும் ஆக்ஸிஜனேற்ற பாதுகாப்பு ஆகியவற்றின் உற்பத்திக்கு இடையிலான சமநிலையில் ஏற்படும் இடையூறு என வரையறுக்கப்படுகிறது. வேறு வார்த்தைகளில் கூறுவதானால், இது ஃப்ரீ ரேடிக்கல்களின் உற்பத்திக்கும், ஆன்டிஆக்ஸிடன்ட்களால் நடுநிலையாக்குவதன் மூலம் தீங்கு விளைவிக்கும் விளைவுகளை எதிர்க்கும் அல்லது நச்சுத்தன்மையாக்கும் உடலின் திறனுக்கும் இடையே உள்ள ஏற்றத்தாழ்வு ஆகும். ஆக்ஸிஜனேற்ற அழுத்தம் உடலில் பல நோய்க்குறியியல் நிலைமைகளுக்கு வழிவகுக்கிறது. நரம்பியக்கடத்தல் நோய்கள், அதாவது பார்கின்சன் நோய், அல்சைமர் நோய், மரபணு மாற்றங்கள், புற்றுநோய்கள், நாள்பட்ட சோர்வு நோய்க்குறி, உடையக்கூடிய X நோய்க்குறி, இதயம் மற்றும் இரத்த நாளக் கோளாறுகள், பெருந்தமனி தடிப்பு, இதய செயலிழப்பு, மாரடைப்பு மற்றும் அழற்சி நோய்கள் ஆகியவை இதில் அடங்கும். ஆக்ஸிஜனேற்றம் பல சூழ்நிலைகளில் நிகழ்கிறது:
செல்கள் ஆற்றலை உருவாக்க குளுக்கோஸைப் பயன்படுத்துகின்றன
நோயெதிர்ப்பு அமைப்பு பாக்டீரியாவை எதிர்த்துப் போராடுகிறது மற்றும் வீக்கத்தை உருவாக்குகிறது
உடல்கள் மாசுகள், பூச்சிக்கொல்லிகள் மற்றும் சிகரெட் புகை ஆகியவற்றை நச்சு நீக்குகிறது
ஆக்சிஜனேற்றத்தை ஏற்படுத்தக்கூடிய எந்த நேரத்திலும் நமது உடலில் மில்லியன் கணக்கான செயல்முறைகள் நடைபெறுகின்றன. இங்கே சில அறிகுறிகள் உள்ளன:
களைப்பு
நினைவாற்றல் இழப்பு அல்லது மூளை மூடுபனி
தசை மற்றும் அல்லது மூட்டு வலி
நரை முடியுடன் சுருக்கங்கள்
பார்வைக் குறைவு
தலைவலி மற்றும் சத்தத்திற்கு உணர்திறன்
தொற்றுநோய்க்கு சந்தேகம்
கரிம உணவுகளைத் தேர்ந்தெடுப்பது மற்றும் உங்கள் சூழலில் நச்சுகளைத் தவிர்ப்பது பெரிய வித்தியாசத்தை ஏற்படுத்துகிறது. இது, மன அழுத்தத்தைக் குறைப்பதோடு, ஆக்சிஜனேற்றத்தைக் குறைப்பதில் நன்மை பயக்கும்.
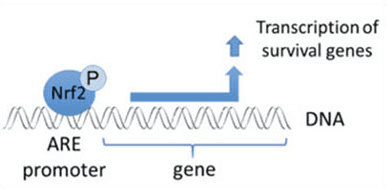
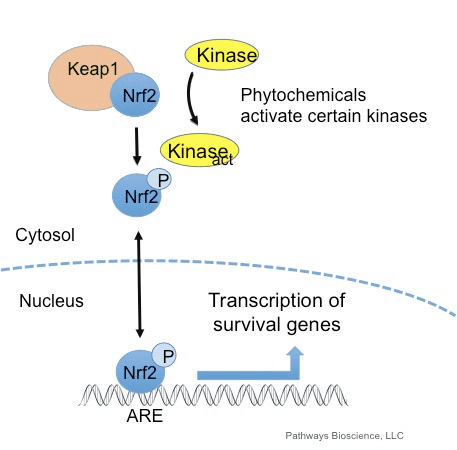
உயிரணுப் பிரிவு, அழற்சி, நோயெதிர்ப்பு செயல்பாடு, தன்னியக்கம் மற்றும் மன அழுத்த பதில் உள்ளிட்ட மனித உடலில் அத்தியாவசிய செயல்முறைகளை ஒழுங்குபடுத்துவதற்காக ஆக்ஸிஜனேற்றங்கள் பொதுவாக கட்டுப்படுத்தப்பட்ட முறையில் உற்பத்தி செய்யப்படுகின்றன. இருப்பினும், இந்த ஆக்ஸிஜனேற்றங்களின் கட்டுப்பாடற்ற உற்பத்தி பங்களிக்க முடியும் விஷத்தன்மை அழுத்தம், இது செல்லுலார் செயல்பாட்டை பாதிக்கலாம், இது நச்சுத்தன்மை, நாள்பட்ட நோய் மற்றும் புற்றுநோயின் வளர்ச்சிக்கு வழிவகுக்கும். மனித உடலின் பாதுகாப்பு ஆக்ஸிஜனேற்ற வழிமுறைகள் ஆக்ஸிஜனேற்றங்களுக்கு செல்லின் பதிலைக் கட்டுப்படுத்தும் முக்கியமான பாதைகளின் வரிசையால் கட்டுப்படுத்தப்படுகின்றன. அணுக்கரு காரணி எரித்ராய்டு 2-தொடர்புடைய காரணி, இல்லையெனில் Nrf2 என அழைக்கப்படுகிறது, இது ஆக்ஸிஜனேற்றங்களுக்கு செல்லுலார் எதிர்ப்பின் வளர்ந்து வரும் சீராக்கி ஆகும். கீழே உள்ள கட்டுரையின் நோக்கம், மைட்டோகாண்ட்ரியல் செயல்பாட்டில் Nrf2 இன் வளர்ந்து வரும் பங்கைப் பற்றி விவாதித்து நிரூபிப்பதாகும்.
சுருக்கம்
டிரான்ஸ்கிரிப்ஷன் காரணி NF-E2 p45-தொடர்புடைய காரணி 2 (Nrf2; மரபணு பெயர் NFE2L2) ஆக்ஸிஜனேற்ற, அழற்சி எதிர்ப்பு மற்றும் நச்சுத்தன்மை என்சைம்கள் உட்பட சைட்டோபிராக்டிவ் புரதங்களின் பல்வேறு நெட்வொர்க்குகளின் மரபணு வெளிப்பாட்டைக் கட்டுப்படுத்துவதன் மூலம் மன அழுத்த சூழ்நிலைகளில் தழுவல் மற்றும் உயிர்வாழ அனுமதிக்கிறது. சேதமடைந்த மேக்ரோமிகுலூல்களை சரிசெய்ய அல்லது அகற்ற உதவும் புரதங்களாக. குளுதாதயோன், தியோரெடாக்சின் மற்றும் NADPH ஆகியவற்றின் உயிரியக்கவியல், பயன்பாடு மற்றும் மீளுருவாக்கம் மற்றும் மைட்டோகாண்ட்ரியா மற்றும் NADPH ஆக்சிடேஸ் மூலம் எதிர்வினை ஆக்ஸிஜன் இனங்களின் உற்பத்தியைக் கட்டுப்படுத்துவதன் மூலம் செல்லுலார் ரெடாக்ஸ் ஹோமியோஸ்டாசிஸை பராமரிப்பதில் Nrf2 முக்கிய பங்கு வகிக்கிறது. ஹோமியோஸ்ட்டிக் நிலைமைகளின் கீழ், Nrf2 மைட்டோகாண்ட்ரியல் சவ்வு திறன், கொழுப்பு அமில ஆக்சிஜனேற்றம், சுவாசத்திற்கான அடி மூலக்கூறுகள் (NADH மற்றும் FADH2/succinate) மற்றும் ATP தொகுப்பு ஆகியவற்றை பாதிக்கிறது. மன அழுத்தம் அல்லது வளர்ச்சிக் காரணி தூண்டுதலின் நிலைமைகளின் கீழ், Nrf2 இன் செயல்பாடானது, மைட்டோகாண்ட்ரியாவில் அதிகரித்த வினைத்திறன் ஆக்ஸிஜன் இனங்கள் உற்பத்தியை எதிர்க்கிறது. கோஆக்டிவேட்டர் 3?, அத்துடன் ப்யூரின் நியூக்ளியோடைடு உயிரியக்கத்தை ஊக்குவிப்பதன் மூலம். இயற்கையாக நிகழும் ஐசோதியோசயனேட் சல்ஃபோராபேன் போன்ற மருந்தியல் Nrf1 ஆக்டிவேட்டர்கள், மைட்டோகாண்ட்ரியல் ஊடுருவல் துளை மற்றும் மைட்டோகாண்ட்ரியல் வீக்கத்தின் ஆக்ஸிஜனேற்ற-மத்தியஸ்த திறப்பைத் தடுக்கின்றன. சுவாரஸ்யமாக, ஒரு செயற்கையான 1-டிஃபெனைல்-2-ட்ரையசோல் கலவை, முதலில் Nrf1,4 ஆக்டிவேட்டராக வடிவமைக்கப்பட்டது, இது மைட்டோபாகியை ஊக்குவிப்பதாகக் கண்டறியப்பட்டது, இதன் மூலம் ஒட்டுமொத்த மைட்டோகாண்ட்ரியல் ஹோமியோஸ்டாசிஸுக்கு பங்களிக்கிறது. எனவே, மைட்டோகாண்ட்ரியாவின் கட்டமைப்பு மற்றும் செயல்பாட்டு ஒருமைப்பாட்டை ஆதரிப்பதில் Nrf1,2,3 முக்கிய பங்கு வகிக்கிறது, மேலும் மன அழுத்த சூழ்நிலைகளில் இந்த பங்கு மிகவும் முக்கியமானது.
முக்கிய வார்த்தைகள்:பயோஎனெர்ஜெடிக்ஸ், சைட்டோபுரோடெக்ஷன், கீப்1, மைட்டோகாண்ட்ரியா, என்ஆர்எஃப்2, ஃப்ரீ ரேடிக்கல்கள்
ஹைலைட்ஸ்
செல்லுலார் ரெடாக்ஸ் ஹோமியோஸ்டாசிஸை பராமரிப்பதில் Nrf2 முக்கிய பங்கு வகிக்கிறது.
Nrf2 மைட்டோகாண்ட்ரியல் சவ்வு திறன் மற்றும் ATP தொகுப்பு ஆகியவற்றை பாதிக்கிறது.
Nrf2 மைட்டோகாண்ட்ரியல் கொழுப்பு அமில ஆக்சிஜனேற்றத்தை பாதிக்கிறது.
Nrf2 மைட்டோகாண்ட்ரியாவின் கட்டமைப்பு மற்றும் செயல்பாட்டு ஒருமைப்பாட்டை ஆதரிக்கிறது.
மைட்டோகாண்ட்ரியல் செயல்பாடு சமரசம் செய்யப்படும்போது Nrf2 ஆக்டிவேட்டர்கள் நன்மை பயக்கும்.
அறிமுகம்
டிரான்ஸ்கிரிப்ஷன் காரணி NF-E2 p45-தொடர்புடைய காரணி 2 (Nrf2; மரபணு பெயர் NFE2L2) பல்வேறு சைட்டோபுரோடெக்டிவ் செயல்பாடுகளுடன் புரதங்களை குறியாக்கம் செய்யும் மரபணுக்களின் நெட்வொர்க்குகளின் வெளிப்பாட்டைக் கட்டுப்படுத்துகிறது. Nrf2 முதன்மையாக புரத நிலைத்தன்மையின் மட்டத்தில் கட்டுப்படுத்தப்படுகிறது. அடிப்படை நிலைமைகளின் கீழ், Nrf2 என்பது ஒரு குறுகிய கால புரதமாகும், இது தொடர்ச்சியான எங்கும் பரவுதல் மற்றும் புரோட்டீசோமால் சிதைவுக்கு உட்பட்டது. Nrf2 இன் சீரழிவுக்கு பங்களிக்கும் மூன்று அறியப்பட்ட ubiquitin ligase அமைப்புகள் உள்ளன. வரலாற்று ரீதியாக, Nrf2 இன் முதல் எதிர்மறை சீராக்கி கண்டுபிடிக்கப்பட்டது Kelch-போன்ற ECH-தொடர்புடைய புரதம் 1 (Keap1) [1], Cullin 3 (Cul3)/Rbx1 ubiquitin ligase [2], [3], [ 4]. Keap1, எங்கும் பரவுதல் மற்றும் புரோட்டீசோமால் சிதைவுக்கு Nrf2 ஐ குறிவைக்க மிகவும் திறமையான சுழற்சி பொறிமுறையைப் பயன்படுத்துகிறது, இதன் போது Keap1 தொடர்ந்து மீளுருவாக்கம் செய்யப்படுகிறது, இது சுழற்சியை தொடர அனுமதிக்கிறது (படம் 1A) [5]. கிளைகோஜன் சின்தேஸ் கைனேஸ் (GSK)2/?-TrCP-சார்ந்த Cul3-அடிப்படையிலான ubiquitin ligase [1], [6] மூலம் Nrf7 சீரழிவுக்கு உட்பட்டது. மிக சமீபத்தில், எண்டோபிளாஸ்மிக் ரெட்டிகுலம் அழுத்தத்தின் போது, E2 ubiquitin ligase Hrd3 [1] மூலம் மத்தியஸ்தம் செய்யப்பட்ட செயல்பாட்டில் Nrf8 எங்கும் பரவுகிறது மற்றும் சிதைகிறது என்று தெரிவிக்கப்பட்டது.
படம் 1 Nrf1 இன் Keap2-மத்தியஸ்த சிதைவுக்கான சுழற்சி தொடர் பிணைப்பு மற்றும் மீளுருவாக்கம் மாதிரி. (A) Nrf2 ஒரு இலவச Keap1 டைமருடன் வரிசையாக பிணைக்கிறது: முதலில் அதன் உயர்-இணைப்பு ETGE (சிவப்பு குச்சிகள்) பிணைப்பு டொமைன் மற்றும் அதன் குறைந்த-இணைப்பு DLG (கருப்பு குச்சிகள்) பிணைப்பு டொமைன் மூலம். புரத வளாகத்தின் இந்த இணக்கத்தில், Nrf2 எங்கும் பரவுகிறது மற்றும் புரோட்டீசோமால் சிதைவுக்கு இலக்காகிறது. இலவச Keap1 ஆனது, புதிதாக மொழிபெயர்க்கப்பட்ட Nrf2 உடன் இணைக்கப்பட்டு, மீண்டும் மீண்டும் தொடங்குகிறது.(B) தூண்டிகள் (வெள்ளை வைரங்கள்) Keap1 இன் சென்சார் சிஸ்டைன்களுடன் (நீல குச்சிகள்) வினைபுரிகின்றன, இது இணக்கமான மாற்றம் மற்றும் பலவீனமான அடி மூலக்கூறு செயல்பாட்டிற்கு வழிவகுக்கிறது. Free Keap1 மீண்டும் உருவாக்கப்படவில்லை, மேலும் புதிதாக ஒருங்கிணைக்கப்பட்ட Nrf2 குவிந்து கருவாக மாறுகிறது.
எபிக்விடின் லிகேஸ் சப்ஸ்ட்ரேட் அடாப்டர் புரோட்டீனாக சேவை செய்வதோடு, கீப்1 என்பது Nrf2 (இன்ட்யூசர்கள் என அழைக்கப்படுகிறது) [9] சிறிய-மூலக்கூறு செயல்பாட்டாளர்களின் பரந்த வரிசைக்கான சென்சார் ஆகும். Keap1 [2], [1] க்குள் குறிப்பிட்ட சிஸ்டைன் எச்சங்களை வேதியியல் முறையில் மாற்றியமைப்பதன் மூலம் அல்லது Keap10:Nrf11 பிணைப்பு இடைமுகத்தை [1], [2] நேரடியாக சீர்குலைப்பதன் மூலம் தூண்டிகள் Nrf12 இன் Keap13-மத்தியஸ்த சிதைவின் சுழற்சியைத் தடுக்கின்றன. இதன் விளைவாக, Nrf2 சிதைவடையவில்லை, மேலும் டிரான்ஸ்கிரிப்ஷன் காரணி குவிந்து கருவுக்கு இடமாற்றம் செய்யப்படுகிறது (படம் 1B), அங்கு அது ஒரு சிறிய மாஃப் புரதத்துடன் ஒரு ஹீட்டோரோடைமரை உருவாக்குகிறது; ஆக்ஸிஜனேற்ற-பதிலளிப்பு கூறுகளுடன் பிணைக்கிறது, அதன் இலக்கு மரபணுக்களின் அப்ஸ்ட்ரீம் ஒழுங்குமுறை பகுதிகள்; மற்றும் டிரான்ஸ்கிரிப்ஷனை துவக்குகிறது [14], [15], [16]. Nrf2 இலக்குகளின் பேட்டரி பல்வேறு சைட்டோபுரோடெக்டிவ் செயல்பாடுகளைக் கொண்ட புரதங்களைக் கொண்டுள்ளது, இதில் ஜீனோபயாடிக் வளர்சிதை மாற்றத்தின் நொதிகள், ஆக்ஸிஜனேற்ற மற்றும் அழற்சி எதிர்ப்பு செயல்பாடுகளைக் கொண்ட புரதங்கள் மற்றும் புரோட்டீசோமல் துணைக்குழுக்கள், அத்துடன் செல்லுலார் ரெடாக்ஸ் ஹோமியோஸ்டாசிஸைக் கட்டுப்படுத்தும் மற்றும் இடைநிலை வளர்சிதை மாற்றத்தில் பங்கேற்கும் புரதங்கள் ஆகியவை அடங்கும்.
செல்லுலார் ரெடாக்ஸ் ஹோமியோஸ்டாசிஸின் முதன்மை சீராக்கியாக Nrf2 இன் செயல்பாடு பரவலாக அங்கீகரிக்கப்பட்டுள்ளது. ?-குளூட்டமைல் சிஸ்டைன் லிகேஸின் வினையூக்கி மற்றும் ஒழுங்குமுறை துணைப்பிரிவுகள் இரண்டின் மரபணு வெளிப்பாடு, குறைக்கப்பட்ட குளுதாதயோனின் (ஜிஎஸ்ஹெச்) உயிரியக்கத் தொகுப்பின் விகித-கட்டுப்படுத்தும் படியை ஊக்குவிக்கும் நொதியானது நேரடியாக Nrf2 ஆல் கட்டுப்படுத்தப்படுகிறது [17]. சிஸ்டைனை கலங்களில் இறக்குமதி செய்யும் சிஸ்டம் xc-யின் xCT துணைக்குழுவும் Nrf2 [18] இன் நேரடி டிரான்ஸ்கிரிப்ஷனல் இலக்காகும். கலத்தில், சிஸ்டைன் சிஸ்டைனாக மாற்றப்படுகிறது, இது ஜிஎஸ்ஹெச் இன் உயிரியக்கத்தின் முன்னோடியாகும். GSH உயிரியக்கத் தொகுப்பில் அதன் பங்கிற்கு கூடுதலாக, Nrf2 குளுதாதயோன் ரிடக்டேஸ் 1 [19], [20] இன் ஒருங்கிணைந்த டிரான்ஸ்கிரிப்ஷனல் ஒழுங்குமுறை மூலம் குளுதாதயோனை அதன் குறைக்கப்பட்ட நிலையில் பராமரிப்பதற்கான வழிமுறைகளை வழங்குகிறது, இது NADPH இலிருந்து சமமானவற்றைக் குறைத்து GSH ஆக ஆக்ஸிஜனேற்றப்பட்ட குளுதாதயோனைக் குறைக்கிறது. . தேவையான NADPH ஆனது நான்கு முக்கிய NADPH-உருவாக்கும் நொதிகளால் வழங்கப்படுகிறது, மாலிக் என்சைம் 1 (ME1), ஐசோசிட்ரேட் டீஹைட்ரோஜினேஸ் 1 (IDH1), குளுக்கோஸ்-6-பாஸ்பேட் டீஹைட்ரோஜினேஸ் (G6PD), மற்றும் 6-பாஸ்போகுளுகோனேட் டீஹைட்ரோஜினேஸ் (PGD), இவை அனைத்தும் டிரான்ஸ்கிரிப்ஷனலாக Nrf2 (படம் 2) [21], [22], [23], [24] மூலம் கட்டுப்படுத்தப்பட்டது. சுவாரஸ்யமாக, ஆல்டிஹைட் டீஹைட்ரோஜினேஸின் சைட்டோசோலிக், மைக்ரோசோமல் மற்றும் மைட்டோகாண்ட்ரியல் வடிவங்களின் தூண்டக்கூடிய மரபணு வெளிப்பாட்டையும் Nrf2 ஒழுங்குபடுத்துகிறது [25], இது NAD(P)+ ஐ இணை காரணியாகப் பயன்படுத்துகிறது, NAD(P)H ஐ உருவாக்குகிறது. உண்மையில், Nrf2-நாக் அவுட் (Nrf2-KO) எலிகளிலிருந்து தனிமைப்படுத்தப்பட்ட கரு இழைமங்களில் NADPH மற்றும் NADPH/NADP+ விகிதங்கள் அவற்றின் காட்டு-வகை (WT) சகாக்களின் செல்களுடன் ஒப்பிடும்போது குறைவாகவே உள்ளன, மேலும் Nrf2 நாக் டவுனில் NADPH அளவுகள் குறையும். அமைப்புரீதியாக செயல்படும் Nrf2 [26] உடன் புற்றுநோய் செல் கோடுகள். எதிர்பார்த்தபடி, Nrf2 சீர்குலைந்த கலங்களில் GSH இன் அளவு குறைவாக உள்ளது; மாறாக, மரபணு அல்லது மருந்தியல் வழிமுறைகளால் Nrf2 செயல்படுத்தல் GSH முறைப்படுத்தலுக்கு வழிவகுக்கிறது [27], [28], [29]. முக்கியமாக, Nrf2 தியோரெடாக்சின் [30], [31], [32], தியோரெடாக்சின் ரிடக்டேஸ் 1 [28], [29], [32], [33], மற்றும் சல்ஃபைர்டாக்சின் [34] ஆகியவற்றின் மரபணு வெளிப்பாட்டையும் ஒழுங்குபடுத்துகிறது. ஆக்ஸிஜனேற்றப்பட்ட புரத தியோல்களைக் குறைப்பதற்காக.
படம் 2 வேகமாகப் பெருகும் உயிரணுக்களின் வளர்சிதை மாற்றத்தில் Nrf2 இன் பங்கு. Nrf2 என்பது ஆக்ஸிஜனேற்ற கை [அதாவது, குளுக்கோஸ்-6-பாஸ்பேட் டீஹைட்ரோஜினேஸ் (G6PD) மற்றும் 6-பாஸ்போகுளோகோனேட் டீஹைட்ரோஜினேஸ் (PGD)] மற்றும் ஆக்சிஜனேற்றம் இல்லாத கை [அதாவது, ட்ரான்சால்டோலேஸ் 1 (TALDO1) மற்றும் டிரான்ஸ்கெட்டோலாஸ் ஆகிய இரண்டிலும் உள்ள மரபணு குறியாக்க என்சைம்களின் நேர்மறை சீராக்கி ஆகும். TKT)] பென்டோஸ் பாஸ்பேட் பாதை. G6PD மற்றும் PGD ஆகியவை NADPH ஐ உருவாக்குகின்றன. Nrf2 மற்ற இரண்டு NADPH-உருவாக்கும் என்சைம்களான மாலிக் என்சைம் 1 (ME1) மற்றும் ஐசோசிட்ரேட் டீஹைட்ரோஜினேஸ் 1 (IDH1) ஆகியவற்றின் மரபணு வெளிப்பாட்டையும் ஒழுங்குபடுத்துகிறது. டி நோவோ பியூரின் பயோசிந்தெடிக் பாதையில் நுழைவதை ஊக்குவிக்கும் பாஸ்போரிபோசில் பைரோபாஸ்பேட் அமிடோட்ரான்ஸ்ஃபெரேஸின் (பிபிஏடி) மரபணு வெளிப்பாடு, மெத்திலீனெட்ரஹைட்ரோஃபோலேட் டீஹைட்ரோஜினேஸ் 2 (எம்டிஎச்எஃப்டி2) இன் முக்கியப் பாத்திரமான என்சியோமியோன்ட்ரியல் இன் வெளிப்பாடாக Nrf2 ஆல் நேர்மறையாகக் கட்டுப்படுத்தப்படுகிறது. டி நோவோ பியூரின் உயிரியக்கத்திற்கு ஒரு கார்பன் அலகுகளை வழங்குகிறது. Pyruvate kinase (PK) ஆனது Nrf2 ஆல் எதிர்மறையாகக் கட்டுப்படுத்தப்படுகிறது மற்றும் கிளைகோலைடிக் இடைநிலைகளின் உருவாக்கத்திற்கும், G6PD உடன் இணைந்து, பென்டோஸ் பாஸ்பேட் பாதை மற்றும் நியூக்ளிக் அமிலங்கள், அமினோ அமிலங்கள் மற்றும் பாஸ்போலிப்பிட்களின் தொகுப்பு ஆகியவற்றின் மூலம் வளர்சிதை மாற்றத்தை சேனலுக்கும் சாதகமாக இருக்கும் என்று எதிர்பார்க்கப்படுகிறது. Nrf2 ஆனது ATP-சிட்ரேட் லைஸின் (CL) மரபணு வெளிப்பாட்டை எதிர்மறையாக ஒழுங்குபடுத்துகிறது, இது மைட்டோகாண்ட்ரியல் பயன்பாட்டிற்கான சிட்ரேட்டின் கிடைக்கும் தன்மையை அதிகரிக்கலாம் அல்லது IDH1 க்கு (ஐசோசிட்ரேட் மூலம்). சிவப்பு மற்றும் நீலம் முறையே நேர்மறை மற்றும் எதிர்மறை கட்டுப்பாட்டைக் குறிக்கின்றன. மைட்டோகாண்ட்ரியன் சாம்பல் நிறத்தில் காட்டப்பட்டுள்ளது. வளர்சிதை மாற்ற சுருக்கங்கள்: G-6-P, குளுக்கோஸ் 6-பாஸ்பேட்; F-6-P, பிரக்டோஸ் 6-பாஸ்பேட்; F-1,6-BP, பிரக்டோஸ் 1,6-பிஸ்பாஸ்பேட்; GA-3-P, கிளைசெரால்டிஹைட் 3-பாஸ்பேட்; 3-PG, 3-பாஸ்போகிளிசரேட்; PEP, phosphoenolpyruvate; 6-பி-ஜிஎல், 6-பாஸ்போகுளோகோனோலாக்டோன்; 6-PG, 6-பாஸ்போகுளுகோனேட்; R-5-P, ரிபுலோஸ் 5-பாஸ்பேட்; PRPP, 5-பாஸ்போரிபோசில்-?-1-பைரோபாஸ்பேட்; THF, டெட்ராஹைட்ரோஃபோலேட்; IMP, இனோசின் மோனோபாஸ்பேட்; AMP, அடினோசின் மோனோபாஸ்பேட்; ஜிஎம்பி, குவானோசின் மோனோபாஸ்பேட்.
செல்லுலார் ரெடாக்ஸ் ஹோமியோஸ்டாசிஸின் முதன்மை சீராக்கியாக Nrf2 இன் முக்கிய பங்கைக் கருத்தில் கொண்டு, WT செல்களுடன் ஒப்பிடும்போது, Nrf2 சீர்குலைந்த கலங்களில் எதிர்வினை ஆக்ஸிஜன் இனங்களின் (ROS) அளவுகள் அதிகமாக இருப்பதில் ஆச்சரியமில்லை (Nrf2-KO) [35]. ஆக்ஸிஜனேற்ற அழுத்தத்தை ஏற்படுத்தும் முகவர்களுடனான சவாலின் மீது இந்த வேறுபாடு குறிப்பாக வேலைநிறுத்தம் செய்கிறது. மேலும், Nrf2 இல் குறைபாடுள்ள செல்கள் பல்வேறு வகையான ஆக்ஸிஜனேற்றிகளின் நச்சுத்தன்மைக்கு அதிக உணர்திறன் கொண்டவை மற்றும் Nrf2 தூண்டிகளால் பாதுகாக்க முடியாது, அதே நிலைமைகளின் கீழ், WT செல்களுக்கு [29], [36] திறமையான மற்றும் நீடித்த பாதுகாப்பை வழங்குகிறது. , [37]. ஒட்டுமொத்த செல்லுலார் ரெடாக்ஸ் ஹோமியோஸ்டாசிஸுடன் கூடுதலாக, மைட்டோகாண்ட்ரியல் ரெடாக்ஸ் ஹோமியோஸ்டாசிஸின் பராமரிப்பிற்கும் Nrf2 முக்கியமானது. எனவே, WT உடன் ஒப்பிடும்போது, Keap1-KO இல் மொத்த மைட்டோகாண்ட்ரியல் NADH குளம் கணிசமாக அதிகரித்துள்ளது மற்றும் Nrf2-KO கலங்களில் வியத்தகு அளவில் குறைந்துள்ளது [35].
லைவ் செல் இமேஜிங்கைப் பயன்படுத்தி, முதன்மை குளோனியூரோனல் கலாச்சாரங்கள் மற்றும் WT, Nrf2-KO அல்லது Keap1-knockdown (Keap1-KD) எலிகளிலிருந்து தனிமைப்படுத்தப்பட்ட மூளை திசு துண்டுகள் [38] ஆகியவற்றில் ROS உற்பத்தியின் விகிதங்களை நாங்கள் சமீபத்தில் கண்காணித்தோம். எதிர்பார்த்தபடி, Nrf2-KO செல்கள் மற்றும் திசுக்களில் அவற்றின் WT சகாக்களுடன் ஒப்பிடும்போது ROS உற்பத்தி விகிதம் வேகமாக இருந்தது. எவ்வாறாயினும், WT உடன் ஒப்பிடும்போது, Keap1-KD செல்கள் ROS உற்பத்தியின் அதிக விகிதங்களைக் கொண்டுள்ளன என்பதை நாங்கள் எதிர்பாராத அவதானிப்பை மேற்கொண்டோம், இருப்பினும் WT மற்றும் Keap1-KD மரபணு வகைகளுக்கு இடையேயான வித்தியாசத்தின் அளவு WT மற்றும் Nrf2-KO இடையே இருந்ததை விட சிறியதாக இருந்தது. . மூளை நோயியலில் உட்படுத்தப்பட்ட இரண்டு NADPH ஆக்சிடேஸ் (NOX) ஐசோஃபார்ம்களின் வினையூக்க துணைக்குழுகளான NOX2 மற்றும் NOX4 இன் mRNA அளவை நாங்கள் பகுப்பாய்வு செய்தோம், மேலும் Nrf2 குறைபாடு நிலைமைகளின் கீழ் NOX2 வியத்தகு அளவில் அதிகரித்திருப்பதைக் கண்டறிந்தோம், அதேசமயம் Nrf4 போது NOX2 கட்டுப்படுத்தப்படுகிறது. சிறிய அளவில் இருந்தாலும், அமைப்புரீதியாக செயல்படுத்தப்படுகிறது. அளவுரீதியாக, பிறழ்ந்த எலிகளிலிருந்து செல்கள் மற்றும் திசுக்களில் உள்ள ஒழுங்குபடுத்தலின் அளவு ROS உற்பத்தியில் தொடர்புடைய அதிகரிப்புக்கு இணையாக உள்ளது [38]. சுவாரஸ்யமாக, Nrf2 NADPH ஆக்சிடேஸை ஒழுங்குபடுத்துவது மட்டுமல்லாமல், NADPH ஆக்சிடேஸால் உற்பத்தி செய்யப்படும் ROS ஆனது நுரையீரல் எபிடெலியல் செல்கள் மற்றும் கார்டியோமயோசைட்டுகள் [2], [39] இல் காட்டப்பட்டுள்ளபடி Nrf40 ஐ செயல்படுத்த முடியும். மேலும், ஒரு மிக சமீபத்திய ஆய்வு Nrf2 இன் NADPH ஆக்சிடேஸ்-சார்ந்த செயல்படுத்தல் மைட்டோகாண்ட்ரியல் சேதம் மற்றும் நாள்பட்ட அழுத்த சுமையின் போது இதயத்தில் உயிரணு இறப்பிற்கு எதிராக பாதுகாப்பதற்கான ஒரு முக்கியமான எண்டோஜெனஸ் பொறிமுறையை உருவாக்குகிறது என்பதை நிரூபித்துள்ளது [41].
NADPH ஆக்சிடேஸின் வினையூக்க செயல்பாட்டிற்கு கூடுதலாக, மைட்டோகாண்ட்ரியல் சுவாசம் என்பது ROS இன் மற்றொரு முக்கிய உள்செல்லுலார் மூலமாகும். மைட்டோகாண்ட்ரியா-குறிப்பிட்ட ஆய்வு MitoSOX ஐப் பயன்படுத்துவதன் மூலம், மைட்டோகாண்ட்ரியாவின் ROS இன் பங்களிப்பை மைட்டோகாண்ட்ரியாவின் ஒட்டுமொத்த ROS உற்பத்தியில் முதன்மையான glioneuronal coculturonal cocultural cocultural உற்பத்தியில் ஆய்வு செய்துள்ளோம். WT, Nrf2-KO, அல்லது Keap1-KD எலிகளிலிருந்து [38]. எதிர்பார்த்தபடி, WT ஐ விட Nrf2-KO செல்கள் மைட்டோகாண்ட்ரியல் ROS உற்பத்தியின் அதிக விகிதங்களைக் கொண்டிருந்தன. ஒட்டுமொத்த ROS உற்பத்திக்கான கண்டுபிடிப்புகளுடன் உடன்பாட்டில், WT செல்களுடன் ஒப்பிடும்போது Keap1-KD இல் மைட்டோகாண்ட்ரியல் ROS உற்பத்தியின் விகிதங்களும் அதிகமாக இருந்தன. முக்கியமாக, ரோட்டெனோனுடன் காம்ப்ளக்ஸ் I ஐ தடுப்பது WT மற்றும் Keap1-KD செல்கள் இரண்டிலும் மைட்டோகாண்ட்ரியல் ROS உற்பத்தியில் வியத்தகு அதிகரிப்பை ஏற்படுத்தியது, ஆனால் Nrf2-KO செல்களில் எந்த விளைவையும் ஏற்படுத்தவில்லை. பைருவேட்டைச் சேர்த்த பிறகு WT செல்களில் மைட்டோகாண்ட்ரியல் ROS உற்பத்தியில் எதிர்பார்க்கப்படும் அதிகரிப்புக்கு மாறாக (NADH இன் கிடைக்கும் தன்மையை அதிகரிக்க, மைட்டோகாண்ட்ரியல் சவ்வு திறனை அதிகரிக்க, மற்றும் சுவாசத்தை இயல்பாக்க), ROS இன் உற்பத்தி Nrf2-KO செல்களில் குறைந்தது. ஒன்றாக, இந்த கண்டுபிடிப்புகள் Nrf2 இல்லாத நிலையில்: (i) சிக்கலான I இன் செயல்பாடு பலவீனமடைகிறது, (ii) சிக்கலான I இன் பலவீனமான செயல்பாடு அடி மூலக்கூறுகளின் வரம்பு காரணமாக உள்ளது, மற்றும் (iii) சிக்கலான செயல்பாடு பலவீனமடைகிறது. மைட்டோகாண்ட்ரியல் ROS உற்பத்தி அதிகரிப்பதற்கு நான் முக்கிய காரணங்களில் ஒன்றாகும், இது சிக்கலான II இலிருந்து எதிர் எலக்ட்ரான் ஓட்டம் காரணமாக இருக்கலாம்.
Nrf2 மைட்டோகாண்ட்ரியல் சவ்வு திறன் மற்றும் சுவாசத்தை பாதிக்கிறது
மைட்டோகாண்ட்ரியல் சவ்வு திறன் (??m) என்பது மைட்டோகாண்ட்ரியல் ஆரோக்கியம் மற்றும் உயிரணுவின் வளர்சிதை மாற்ற நிலை ஆகியவற்றின் உலகளாவிய குறிகாட்டியாகும். ஒரு ஆரோக்கியமான கலத்தில், ??m மைட்டோகாண்ட்ரியல் சுவாச சங்கிலியால் பராமரிக்கப்படுகிறது. சுவாரஸ்யமாக, ஈஸ்ட்ரோஜன் ஏற்பி-எதிர்மறை மனித மார்பக எபிடெலியல் MCF10A செல் லைனில் கலாச்சார அடிப்படையிலான புரோட்டியோமிக்ஸ் ஆய்வில் அமினோ அமிலங்களைக் கொண்ட ஒரு நிலையான ஐசோடோபிக் லேபிளிங், மைட்டோகாண்ட்ரியல் எலக்ட்ரான் போக்குவரத்து சங்கிலி கூறு NDUFA4 NDUFA2 மருந்தியல் செயல்பாட்டின் மூலம் கட்டுப்படுத்தப்படுகிறது என்பதைக் காட்டுகிறது. அதேசமயம் Nrf2 (Keap1 நாக் டவுன் மூலம்) மரபியல் முறைப்படுத்தல் சைட்டோக்ரோம் c ஆக்சிடேஸ் துணை அலகுகளான COX2 மற்றும் COX4I1 [42] ஆகியவற்றைக் குறைக்க வழிவகுக்கிறது. இரு பரிமாண ஜெல் எலக்ட்ரோபோரேசிஸ் மற்றும் மேட்ரிக்ஸ்-அசிஸ்டெட் லேசர் டிசார்ப்ஷன்/அயனியாக்கம் மாஸ் ஸ்பெக்ட்ரோமெட்ரி ஆகியவற்றைப் பயன்படுத்தி கல்லீரல் புரோட்டியோமின் ஆய்வில், Nrf2 ATP சின்தேஸ் சப்யூனிட்டின் வெளிப்பாட்டைக் கட்டுப்படுத்துகிறது என்பதைக் கண்டறிந்துள்ளது. [43]. கூடுதலாக, மைட்டோகாண்ட்ரியல் புரதம் DJ-1, காம்ப்ளக்ஸ் I [44] இன் செயல்பாட்டைப் பராமரிப்பதில் பங்கு வகிக்கிறது, Nrf2 [45], [46] ஐ உறுதிப்படுத்துகிறது, இருப்பினும் மருந்தியல் அல்லது மரபணு செயல்பாட்டின் நரம்பியல் விளைவுகள் Nrf2 இன் DJ-1 [47] இலிருந்து சுயாதீனமானது. இருப்பினும், மைட்டோகாண்ட்ரியல் செயல்பாட்டிற்கான இந்த அவதானிப்புகளின் விளைவுகள் ஆராயப்படவில்லை.
Nrf2 குறைபாட்டின் கீழ் சிக்கலான I இன் பலவீனமான செயல்பாட்டின் உடன்படிக்கையில், Nrf2-KO மவுஸ் எம்ப்ரியோனிக் ஃபைப்ரோபிளாஸ்ட்கள் (MEFகள்) மற்றும் வளர்ப்பு முதன்மை கிளியோனியூரோனல் செல்கள் அவற்றின் WT உடன் ஒப்பிடுகையில் அடித்தள ??m குறைவாக உள்ளது (படம். 3, இன்செட்) [35]. இதற்கு நேர்மாறாக, Nrf2 மரபணு ரீதியாக அமைப்புரீதியாக ஒழுங்குபடுத்தப்படும் போது அடித்தளம் ??m அதிகமாக இருக்கும் (Keap1 இன் நாக் டவுன் அல்லது நாக் அவுட் மூலம்). மரபணு வகைகளில் ??m இல் உள்ள இந்த வேறுபாடுகள் Nrf2 இன் செயல்பாட்டால் சுவாசம் பாதிக்கப்படுகிறது என்பதைக் குறிக்கிறது. உண்மையில், அடித்தள நிலையில் உள்ள ஆக்ஸிஜன் நுகர்வு மதிப்பீடு, WT உடன் ஒப்பிடும்போது, Nrf2-KO மற்றும் Keap1-KO MEF களில் ஆக்ஸிஜன் நுகர்வு முறையே ~50 மற்றும் ~35% குறைவாக உள்ளது என்பதை வெளிப்படுத்தியுள்ளது.
படம் 3 Nrf2 குறைபாடு நிலைமைகளின் கீழ் சமரசம் செய்யப்பட்ட மைட்டோகாண்ட்ரியல் செயல்பாட்டிற்கான முன்மொழியப்பட்ட வழிமுறை. (1) ME1, IDH1, G6PD மற்றும் PGD இன் குறைந்த அளவு NADPH அளவைக் குறைக்கிறது. (2) GSH இன் அளவும் குறைவாக உள்ளது. (3) ME1 இன் குறைந்த செயல்பாடு மைட்டோகாண்ட்ரியாவுக்குள் நுழையும் பைருவேட்டின் குளத்தை குறைக்கலாம். (4) NADH இன் உருவாக்கம் மெதுவாக உள்ளது, இது சிக்கலான I இன் பலவீனமான செயல்பாடு மற்றும் அதிகரித்த மைட்டோகாண்ட்ரியல் ROS உற்பத்திக்கு வழிவகுக்கிறது. (5) மைட்டோகாண்ட்ரியல் புரதங்களில் FAD ஐ FADH2 ஆகக் குறைப்பதும் குறைக்கப்படுகிறது, இது FADH2 இலிருந்து UbQ மற்றும் சிக்கலான III ஆக எலக்ட்ரான் ஓட்டத்தைக் குறைக்கிறது. (6) UbQH2 இன் மெதுவான உருவாக்கம் சக்சினேட் டீஹைட்ரோஜினேஸின் நொதி செயல்பாட்டைக் குறைக்கலாம். (7) ROS இன் அதிகரித்த அளவுகள் சிக்கலான II இன் செயல்பாட்டை மேலும் தடுக்கலாம். (8) கொழுப்பு அமில ஆக்சிஜனேற்றத்தின் குறைந்த செயல்திறன் மைட்டோகாண்ட்ரியல் சுவாசத்திற்கான அடி மூலக்கூறு கிடைப்பதில் குறைவுக்கு பங்களிக்கிறது. (9) ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷனில் ஏடிபி உற்பத்தி குறைவதற்கான ஈடுசெய்யும் பொறிமுறையாக கிளைகோலிசிஸ் மேம்படுத்தப்படுகிறது. (10) ATP சின்தேஸ் ??m பராமரிக்க தலைகீழாக செயல்படுகிறது. சிவப்பு மற்றும் நீலம் முறையே அதிக கட்டுப்பாடு மற்றும் குறைப்பு ஆகியவற்றைக் குறிக்கின்றன. பெட்டிகள் சோதனை ஆதாரங்கள் கிடைப்பதைக் குறிக்கின்றன. பொட்டென்டோமெட்ரிக் ஃப்ளோரசன்ட் ஆய்வு டெட்ராமெதில்ரோடமைன் மெத்தில் எஸ்டர் (TMRM; 2 nM) மூலம் காட்சிப்படுத்தப்பட்ட WT மற்றும் Nrf25-KO கார்டிகல் ஆஸ்ட்ரோசைட்டுகளின் மைட்டோகாண்ட்ரியாவின் படங்களை இன்செட் காட்டுகிறது. அளவுகோல், 20 மீ.
மரபணு வகைகளில் ??m மற்றும் சுவாசத்தில் உள்ள இந்த வேறுபாடுகள் மைட்டோகாண்ட்ரியல் சுவாசத்திற்கான அடி மூலக்கூறுகளின் பயன்பாட்டின் விகிதத்தால் பிரதிபலிக்கப்படுகின்றன. ட்ரைகார்பாக்சிலிக் அமிலம் (TCA) சுழற்சிக்கான அடி மூலக்கூறுகளின் பயன்பாடு (மேலேட்/பைருவேட், இது காம்ப்ளக்ஸ் I அடி மூலக்கூறு NADH இன் உற்பத்தியை அதிகரிக்கிறது) அல்லது காம்ப்ளக்ஸ் II இன் அடி மூலக்கூறான மீதைல் சக்சினேட், WT இரண்டிலும் ??m இல் படிப்படியாக அதிகரிப்பை ஏற்படுத்துகிறது. மற்றும் Keap1-KD நியூரான்கள், ஆனால் அதிகரிப்பு விகிதம் Keap1-KD செல்களில் அதிகமாக உள்ளது. மிக முக்கியமாக, இந்த டிசிஏ சுழற்சி அடி மூலக்கூறுகளுக்கான பதிலின் வடிவங்கள் இரண்டு மரபணு வகைகளுக்கு இடையில் வேறுபடுகின்றன, இதன் மூலம் கீப்1-கேடி செல்களில் அடி மூலக்கூறு சேர்க்கையில் ??மீ வேகத்தில் உயர்வது, ஒரு பீடபூமியை விட விரைவான வீழ்ச்சியைத் தொடர்ந்து, வழக்கத்திற்கு மாறானதாகக் கூறுகிறது. வேகமான அடி மூலக்கூறு நுகர்வு. இந்த கண்டுபிடிப்புகள் WT MEF உடன் ஒப்பிடும்போது Keap50-KO இல் [U-70C1] குளுக்கோஸின் 13-மணிநேர துடிப்புக்குப் பிறகு காணப்பட்ட மாலேட், பைருவேட் மற்றும் சக்சினேட் ஆகியவற்றின் மிகக் குறைந்த (6-1%) அளவுகளுடன் நெருங்கிய உடன்பாட்டில் உள்ளன. செல்கள் [24]. Nrf2-KO நியூரான்களில், பைருவேட் மட்டுமே ??m ஐ அதிகரிக்க முடியும், அதேசமயம் மாலேட் மற்றும் மெத்தில் சுசினேட் லேசான டிப்போலரைசேஷன் ஏற்படுகிறது. மைட்டோகாண்ட்ரியல் அடி மூலக்கூறு உற்பத்தியில் Nrf2 இன் விளைவு மைட்டோகாண்ட்ரியல் செயல்பாட்டை Nrf2 பாதிக்கும் முக்கிய வழிமுறையாகத் தெரிகிறது. மைட்டோகாண்ட்ரியல் NADH ரெடாக்ஸ் இன்டெக்ஸ் (சிக்கலான I மூலம் NADH நுகர்வுக்கும் TCA சுழற்சியில் NADPH உற்பத்திக்கும் இடையே உள்ள சமநிலை) Nrf2-KO செல்களில் அவற்றின் WT சகாக்களுடன் ஒப்பிடுகையில் கணிசமாகக் குறைவாக உள்ளது, மேலும், குளங்களின் மீளுருவாக்கம் விகிதங்கள் சிக்கலான IV (NaCN ஐப் பயன்படுத்துவதன் மூலம்) தடுக்கப்பட்ட பிறகு NADH மற்றும் FADH2 ஆகியவை பிறழ்ந்த உயிரணுக்களில் மெதுவாக இருக்கும்.
முரைன் மூளை மற்றும் கல்லீரலில் இருந்து தனிமைப்படுத்தப்பட்ட மைட்டோகாண்ட்ரியாவில், காம்ப்ளக்ஸ் I அல்லது காம்ப்ளக்ஸ் II க்கு அடி மூலக்கூறுகளை நிரப்புவது, Nrf2 செயல்படுத்தப்படும்போது ஆக்ஸிஜன் நுகர்வு விகிதத்தை மிகவும் வலுவாகவும், Nrf2 பாதிக்கப்படும்போது குறைந்த செயல்திறன் கொண்டதாகவும் அதிகரிக்கிறது [35]. எனவே, WT உடன் ஒப்பிடும்போது Keap1-KD இல் அதிக ஆக்ஸிஜன் நுகர்வு விகிதத்தை மாலேட் தூண்டுகிறது, ஆனால் அதன் விளைவு Nrf2-KO மைட்டோகாண்ட்ரியாவில் பலவீனமாக உள்ளது. இதேபோல், ரோட்டெனோனின் முன்னிலையில் (சிக்கலான I தடுக்கப்படும் போது), WT உடன் ஒப்பிடும்போது, கீப்1-கேடியில் அதிக அளவில் ஆக்சிஜன் நுகர்வை சுசினேட் செயல்படுத்துகிறது, அதேசமயம் Nrf2-KO மைட்டோகாண்ட்ரியாவில் பதில் குறைகிறது. கூடுதலாக, Nrf2-KO முதன்மை நரம்பியல் கலாச்சாரங்கள் மற்றும் எலிகள் சிக்கலான II தடுப்பான்கள் 3-நைட்ரோபிரோபியோனிக் அமிலம் மற்றும் மலோனேட் ஆகியவற்றின் நச்சுத்தன்மைக்கு அதிக உணர்திறன் கொண்டவை, அதேசமயம் Nrf2-அதிகப்படியாக அழுத்தும் ஆஸ்ட்ரோசைட்டுகளின் உள்பகுதி மாற்று அறுவை சிகிச்சையானது பாதுகாப்பானது [48], [49]. இதேபோல், Nrf2-KO எலிகள் அதிக உணர்திறன் கொண்டவை, அதேசமயம் Nrf2 இன் மரபணு அல்லது மருந்தியல் செயலாக்கம் 1-மெத்தில்-4-பீனைல்-1 இல் உள்ள சிக்கலான I இன்ஹிபிட்டர் 4-மெத்தில்-1,2,3,6-ஃபைனில்பைரிடினியம் அயனியால் ஏற்படும் நியூரோடாக்சிசிட்டிக்கு எதிராக பாதுகாப்பு விளைவுகளைக் கொண்டுள்ளது. பார்கின்சன் நோயின் 49-டெட்ராஹைட்ரோபிரைடின் விலங்கு மாதிரி [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61], [XNUMX].
சுவாசக் கட்டுப்பாட்டு விகிதம் (RCR), ஸ்டேட் 3 (ADP-தூண்டப்பட்டது) மற்றும் ஸ்டேட் 4 சுவாசம் (ஏடிபி இல்லை) விகிதம் Nrf2 இல்லாத நிலையில் குறைக்கப்படுகிறது, ஆனால் RCR ஆனது Keap1-KD மற்றும் WT மைட்டோகாண்ட்ரியா [35] இடையே ஒத்ததாக உள்ளது. ]. ஆர்.சி.ஆர் என்பது மைட்டோகாண்ட்ரியல் சுவாச சங்கிலியின் செயல்பாட்டை ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷனுடன் இணைக்கும் அளவைக் குறிப்பதால், கீப்1-கேடி மைட்டோகாண்ட்ரியாவில் அதிக சுவாச விகிதம் ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷனின் இணைப்பின் காரணமாக இல்லை என்பதை இந்த கண்டுபிடிப்பு சுட்டிக்காட்டுகிறது. Nrf2 செயல்படுத்தப்படும்போது ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷன் மிகவும் திறமையானதாக இருக்கும் என்று அது மேலும் தெரிவிக்கிறது. Keap1-KD மைட்டோகாண்ட்ரியாவில் உள்ள அதிக சுவாச விகிதம் மைட்டோகாண்ட்ரியல் ROS உற்பத்தியின் உயர் மட்டங்களுடன் ஒத்துப்போகிறது [38] அதிக சுவாச விகிதங்கள் எலக்ட்ரான் கசிவு அதிகரிக்க வழிவகுக்கும். இருப்பினும், ஆக்ஸிஜனேற்ற அழுத்தத்தின் நிலைமைகளின் கீழ், அதிகரித்த ROS உற்பத்தியானது மைட்டோகாண்ட்ரியல் உள் மென்படலத்தின் புரோட்டான் கடத்துத்திறனை அதிகரிக்கிறது மற்றும் அதன் விளைவாக சூப்பர் ஆக்சைடு [2] உற்பத்தியைக் குறைக்கிறது. மிக சமீபத்தில், லிப்பிட் பெராக்ஸைடேஷன் தயாரிப்பு 3-ஹைட்ராக்ஸி-3-நோனெனல் கார்டியோமயோசைட்டுகளில் UCP62 இன் Nrf4-சார்ந்த மேலோட்டத்தை மத்தியஸ்தம் செய்கிறது என்று காட்டப்பட்டது; இஸ்கிமியா ரிபெர்ஃப்யூஷன் [2] போன்ற ஆக்ஸிஜனேற்ற அழுத்தத்தின் நிலைமைகளின் கீழ் பாதுகாப்பிற்கு இது மிகவும் முக்கியமானதாக இருக்கலாம்.
Nrf2 ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷன் மற்றும் ஏடிபியின் தொகுப்பு ஆகியவற்றின் செயல்திறனை பாதிக்கிறது
மூளை மற்றும் கல்லீரல் மைட்டோகாண்ட்ரியாவில், சுவாசத்தில் Nrf2 இன் தாக்கத்துடன் உடன்படுகையில், Nrf2 குறைபாடு ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷனின் செயல்திறன் குறைவதற்கு வழிவகுக்கிறது (ஏடிபி மற்றும் ஆக்ஸிஜன் விகிதத்தால் மதிப்பிடப்பட்டுள்ளது, இது ATP தொகுப்புக்காக உட்கொள்ளப்படுகிறது), அதேசமயம் Nrf2 செயல்படுத்தல் (Keap1 -KD) எதிர் விளைவைக் கொண்டுள்ளது [35]. WT உடன் ஒப்பிடும்போது, Nrf2 இன் அமைப்புரீதியாக ஒழுங்குபடுத்தப்பட்ட கலங்களில் ATP அளவுகள் கணிசமாக அதிகமாகவும், Nrf2 கீழே தள்ளப்படும்போதும் குறைவாகவும் இருக்கும் [64] அல்லது சீர்குலைந்தால் [35]. மேலும், ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷன் (ஒலிகோமைசின்) அல்லது கிளைகோலிசிஸ் (அயோடோஅசெடிக் அமிலம்) தடுப்பான்களின் பயன்பாடு, செல்கள் ஏடிபியை உருவாக்கும் முறையை Nrf2 மாற்றுகிறது என்பதை வெளிப்படுத்தியுள்ளது. எனவே, WT நியூரான்களில், ஒலிகோமைசின் ஏடிபியில் முழுமையான வீழ்ச்சியை ஏற்படுத்துகிறது மற்றும் அயோடோஅசெடிக் அமிலம் எந்த விளைவையும் ஏற்படுத்தாது. குறிப்பிடத்தக்க வகையில், Nrf2-KO செல்களில், ஒலிகோமைசின் ATP அளவை அதிகரிக்கிறது, அவை மெதுவாக, ஆனால் முழுமையாக, iodoacetic அமிலத்தால் குறைக்கப்படுகின்றன, Nrf2 இல்லாவிட்டாலும், கிளைகோலிசிஸ், ஆனால் ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷன் அல்ல, ATP உற்பத்தியின் முக்கிய ஆதாரம் என்பதைக் குறிக்கிறது. சுவாரஸ்யமாக, கீப்1-கேடி செல்களில் ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷனின் செயல்திறன் அதிகரித்தாலும், ஒலிகோமைசின் சேர்ப்பதால் ஏடிபி அளவுகளில் ~80% குறைகிறது, மேலும் அயோடோஅசெடிக் அமிலம் மேலும் ~20% குறைகிறது. எனவே, Nrf2 குறைபாடு அல்லது அதன் அமைப்புமுறை செயல்படுத்தல் ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷனின் பங்களிப்பைக் குறைக்கிறது மற்றும் ATP இன் தொகுப்புக்கு கிளைகோலிசிஸின் பங்களிப்பை அதிகரிக்கிறது. Nrf2 இல்லாதபோது இந்த விளைவு குறிப்பாக உச்சரிக்கப்படுகிறது மற்றும் நடுத்தர [35] குளுக்கோஸின் இருப்பு மற்றும் கிளைகோலைடிக் இடைநிலைகளின் (G-6-P, F-6-P) அதிகரித்த அளவுகளில் ??m இன் சார்புடன் ஒத்துப்போகிறது. , டைஹைட்ராக்ஸிஅசெட்டோன் பாஸ்பேட், பைருவேட் மற்றும் லாக்டேட்) Nrf2 [24] இன் நாக் டவுனுக்குப் பிறகு.
ஒலிகோமைசின் F1F0-ATPase ஐத் தடுப்பதற்குப் பிறகு ATP அளவுகளில் அதிகரிப்பு, Nrf2 இல்லாத நிலையில், F1F0-ATPase ஆனது ATP சின்தேஸாக அல்லாமல் ATPase ஆகச் செயல்படுகிறது, அதாவது, அது தலைகீழாகச் செயல்படுகிறது என்பதைக் குறிக்கிறது. செயல்பாட்டில் இத்தகைய தலைகீழ் மாற்றமானது, இந்த உறுப்பின் செயல்பாட்டு ஒருமைப்பாட்டிற்கு முக்கியமான ??m ஐ பராமரிக்கும் முயற்சியில் உள் மைட்டோகாண்ட்ரியல் சவ்வு முழுவதும் புரோட்டான்களை பம்ப் செய்ய வேண்டியதன் அவசியத்தை பிரதிபலிக்கிறது. F1F0-ATPase இன் செயல்பாட்டின் தலைகீழ் மாற்றமானது, Nrf2-KO செல்களுக்கு ஒலிகோமைசின் நிர்வாகத்தின் மீது காணப்பட்ட மைட்டோகாண்ட்ரியல் டிப்போலரைசேஷன் மூலம் சாட்சியமளிக்கப்படுகிறது, இது அவற்றின் WT அல்லது Keap1-குறைபாடுள்ள சககளில் நிகழும் ஹைப்பர்போலரைசேஷன் [35] க்கு முற்றிலும் மாறாக உள்ளது. ஒட்டுமொத்தமாக, Nrf2 குறைபாட்டின் கீழ் ATP முதன்மையாக கிளைகோலிசிஸில் உற்பத்தி செய்யப்படுகிறது.
Nrf2 மைட்டோகாண்ட்ரியல் கொழுப்பு அமில ஆக்சிஜனேற்றத்தை மேம்படுத்துகிறது
??m இல் Nrf2 குறைபாட்டின் விளைவு குறிப்பாக செல்கள் குளுக்கோஸ் இல்லாமல் நடுத்தரத்தில் அடைகாக்கப்படும் போது உச்சரிக்கப்படுகிறது, மேலும் WT செல்களுடன் ஒப்பிடும்போது ??m Nrf50-KO இல் ~2% குறைவாக உள்ளது [35]. குளுக்கோஸ் பற்றாக்குறையின் நிலைமைகளின் கீழ், மைட்டோகாண்ட்ரியல் கொழுப்பு அமில ஆக்சிஜனேற்றம் (FAO) என்பது சுவாசம் மற்றும் ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷனுக்கான அடி மூலக்கூறுகளின் முக்கிய வழங்குநராகும், இது Nrf2 FAO ஐ பாதிக்கலாம் என்று பரிந்துரைக்கிறது. உண்மையில், நீண்ட சங்கிலி (C16:0) நிறைவுற்ற கொழுப்பு அமிலம் பால்மிடிக் அமிலம் மற்றும் குறுகிய சங்கிலி (C6:0) ஹெக்ஸானோயிக் அமிலம் ஆகிய இரண்டிற்கும் FAO இன் செயல்திறன், Keap1-KO MEFகள் மற்றும் தனிமைப்படுத்தப்பட்ட இதயம் மற்றும் கல்லீரல் மைட்டோகாண்ட்ரியாவில் அவற்றின் செயல்திறனை விட அதிகமாக உள்ளது. WT சகாக்கள், அதேசமயம் இது Nrf2-KO செல்கள் மற்றும் மைட்டோகாண்ட்ரியாவில் குறைவாக உள்ளது [65]. இந்த விளைவுகள் மனிதர்களுக்கும் மிகவும் பொருத்தமானவை: உண்மையில், TCA சுழற்சியின் செயல்பாட்டுடன் FAO இன் சிறந்த ஒருங்கிணைப்பைக் குறிக்கும் வளர்சிதை மாற்ற மாற்றங்கள், கிளாசிக்கல் Nrf2 ஆக்டிவேட்டர் சல்ஃபோராபேனின் முன்னோடியான குளுக்கோராபனின் நிறைந்த உணவுகளுடன் மனித தலையீட்டு ஆய்வுகளில் நிகழ்கின்றன. 66].
மைட்டோகாண்ட்ரியல் FAO இன் முதல் படியின் போது, ?-கார்பனின் சார்பு ஹைட்ரஜன் ஒரு ஹைட்ரைடாக வெளியேறுகிறது, இது FAD கோஃபாக்டரை FADH2 ஆக குறைக்கிறது, இது எலக்ட்ரான்களை சுவாச சங்கிலியில் உள்ள ubiquinone (UbQ) க்கு மாற்றுகிறது, இறுதியில் ATP உற்பத்திக்கு பங்களிக்கிறது. . குளுக்கோஸ் இல்லாத நிலையில் பால்மிடோயில்கார்னைடைன் மூலம் FAO தூண்டுவது WT மற்றும் Keap1-KO கலங்களில் ATP அளவுகளில் எதிர்பார்க்கப்படும் அதிகரிப்புக்கு காரணமாகிறது, Keap1-KO செல்களில் ATP உயர்வது வேகமாக இருப்பதால், Nrf2-KO இல் ஒரே மாதிரியான சிகிச்சையானது ATP மாற்றங்களை உருவாக்காது. MEFகள் [65]. இந்த சோதனையானது, Nrf2 இல்லாத நிலையில், FAO நசுக்கப்படுகிறது, மேலும், Nrf2 குறைபாடு [35], [64] நிலைமைகளின் கீழ் ATP அளவுகள் குறைவதற்கான காரணங்களில் ஒன்றாக FAOவை அடக்குவதைக் குறிக்கிறது.
குறிப்பிடத்தக்க வகையில், Nrf293 அமைதிப்படுத்தப்பட்ட மனித 2 T செல்கள் CPT1 மற்றும் CPT2[67] இன் குறைந்த வெளிப்பாட்டைக் கொண்டுள்ளன, கார்னைடைன் பால்மிட்டோல்ட்ரான்ஸ்ஃபெரேஸின் (CPT) இரண்டு ஐசோஃபார்ம்கள், மைட்டோகாண்ட்ரியல் FAO இல் உள்ள விகிதத்தைக் கட்டுப்படுத்தும் நொதியாகும். உடன்பாட்டில், WT எலிகளுடன் ஒப்பிடும்போது Cpt1 இன் mRNA அளவுகள் Nrf2-KO இன் கல்லீரல்களில் குறைவாக உள்ளது [68]. CPT ஆனது ஒரு நீண்ட சங்கிலி கொழுப்பு அசைல்-CoA இன் அசைல் குழுவை கோஎன்சைம் A இலிருந்து எல்-கார்னைடைனுக்கு மாற்றுகிறது, இதனால் சைட்டோபிளாஸத்திலிருந்து மைட்டோகாண்ட்ரியாவிற்கு அசைல்கார்னைடைனை இறக்குமதி செய்ய அனுமதிக்கிறது. இது இன்றுவரை ஆய்வு செய்யப்படவில்லை என்றாலும், CPT1 வெளிப்பாட்டின் மீதான டிரான்ஸ்கிரிப்ஷனல் விளைவுகளுக்கு கூடுதலாக, Nrf2 அதன் முக்கிய அலோஸ்டெரிக் தடுப்பானான மலோனைல்-கோஏவின் அளவைக் கட்டுப்படுத்துவதன் மூலம் இந்த நொதியின் செயல்பாட்டையும் பாதிக்கலாம். ஏனென்றால், தற்போது தெளிவற்ற ஒரு பொறிமுறையால், Nrf2 ஸ்டீரோயில் CoA டெசாச்சுரேஸ் (SCD) [69] மற்றும் சிட்ரேட் லைஸ் (CL) [69], [70] ஆகியவற்றின் வெளிப்பாட்டை எதிர்மறையாகக் கட்டுப்படுத்துகிறது. சுவாரஸ்யமாக, SCD இன் நாக் அவுட் அல்லது தடுப்பானது பாஸ்போரிலேஷன் மற்றும் AMP-செயல்படுத்தப்பட்ட புரத கைனேஸ் (AMPK) [71], [72], [73] செயல்படுத்தப்படுவதற்கு வழிவகுக்கிறது, மேலும் இது Nrf2 இல்லாத நிலையில், SCD அளவுகள் என்று ஊகிக்க முடியும். அதிகரிக்கும், இதையொட்டி AMPK செயல்பாட்டைக் குறைக்கும். Nrf2-KO எலிகளின் [68] கல்லீரலில் காணப்பட்ட AMPK இன் குறைக்கப்பட்ட புரத அளவுகளால் இது மேலும் ஒருங்கிணைக்கப்படும் எலிகள் [1]. குறைந்த AMPK செயல்பாட்டின் ஒரு விளைவு, அசிடைல்-CoA கார்பாக்சிலேஸின் (ACC) [74] இன் தடுப்பு பாஸ்போரிலேஷனின் (Ser79 இல்) நிவாரணம் ஆகும், இது Nrf75 இல்லாவிட்டாலும், Nrf2 செயல்படுத்தல் [2] மூலம் குறைக்கப்பட்டதால், இது மேலும் படியெடுத்தல் முறையில் அதிகப்படுத்தப்படலாம். ]. உயர் ACC செயல்பாடு, ACCக்கான அடி மூலக்கூறு அசிடைல்-CoA இன் உற்பத்தியை அதிகரிக்கும் ஒழுங்குபடுத்தப்பட்ட CL வெளிப்பாடுடன் இணைந்து, இறுதியில் ACC தயாரிப்பான malonyl-CoA இன் அளவை அதிகரிக்கலாம். அதிக அளவு மலோனைல்-கோஏ சிபிடியைத் தடுக்கிறது, இதனால் கொழுப்பு அமிலங்கள் மைட்டோகாண்ட்ரியாவுக்குள் கடத்தப்படுவதைக் குறைக்கிறது. இறுதியாக, பிளாஸ்மா மற்றும் மைட்டோகாண்ட்ரியல் சவ்வுகளில் கொழுப்பு அமிலங்களை இறக்குமதி செய்யும் ஒரு இடமாற்றம் CD70 [2] இன் வெளிப்பாட்டை Nrf36 நேர்மறையாக ஒழுங்குபடுத்துகிறது. எனவே, மைட்டோகாண்ட்ரியாவில் நீண்ட சங்கிலி கொழுப்பு அமிலங்களின் இறக்குமதியை ஒழுங்குபடுத்துவதன் மூலம் மைட்டோகாண்ட்ரியல் FAO இன் செயல்திறனை Nrf76 பாதிக்கக்கூடிய ஒரு வழிமுறையாகும்.
நேரடி டிரான்ஸ்கிரிப்ஷனல் ஒழுங்குமுறைக்கு கூடுதலாக, செல்லுலார் ரெடாக்ஸ் வளர்சிதை மாற்றத்தில் அதன் விளைவுகளால் மைட்டோகாண்ட்ரியல் FAO இன் செயல்திறனை Nrf2 மாற்றலாம். Nrf2 செயல்பாடு குறைவாகவோ அல்லது இல்லாமலோ இருக்கும்போது, செல்லுலார் ரெடாக்ஸ் நிலையை ஆக்ஸிஜனேற்ற நிலைக்கு மாற்றும் நிலைமைகள் இருக்கும் போது இது மிகவும் பொருத்தமானதாக இருக்கலாம். உண்மையில், பல FAO என்சைம்கள் ரெடாக்ஸ் மாற்றங்களுக்கு உணர்திறன் கொண்டவையாக அடையாளம் காணப்பட்டுள்ளன. அத்தகைய நொதிகளில் ஒன்று மிக நீண்ட சங்கிலி அசைல்-கோஏ டிஹைட்ரோஜினேஸ் (விஎல்சிஏடி) ஆகும், இது மனித திசுக்களில் பால்மிடோயில்-கோஏ டீஹைட்ரஜனேற்றம் செயல்பாட்டிற்கு 80% க்கும் அதிகமாக பங்களிக்கிறது [77]. சுவாரஸ்யமாக, ஹர்ட் மற்றும் பலர். [78] VLCAD ஆனது சிஸ்டைன் எச்சங்களைக் கொண்டிருப்பதைக் காட்டுகிறது, அவை H2O2 க்கு தனிமைப்படுத்தப்பட்ட எலி இதய மைட்டோகாண்ட்ரியாவின் வெளிப்பாட்டின் போது அவற்றின் ரெடாக்ஸ் நிலையை கணிசமாக மாற்றும். கூடுதலாக, Cys238 இல் உள்ள Murine hepatic VLCAD இன் S-நைட்ரோசைலேஷன் நொதியின் வினையூக்க செயல்திறனை மேம்படுத்துகிறது [79], மேலும் அதே சிஸ்டைனின் ஆக்சிஜனேற்றம் எதிர் விளைவைக் கொண்டிருக்கலாம், இறுதியில் மைட்டோகாண்ட்ரியல் FAO இன் செயல்திறனைக் குறைக்கலாம். WT, Nrf2-KO, அல்லது Keap1-KO MEFகள் [65] ஆகியவற்றில் VLCAD இன் வெளிப்பாடு நிலைகள் கணிசமாக வேறுபடவில்லை என்றாலும், Nrf2 இல்லாத நிலையில் VLCAD இன் என்சைம் செயல்பாடு அதிக அளவுகள் காரணமாக குறைவாக இருக்கலாம். ROS இன்.
இந்த அனைத்து கண்டுபிடிப்புகளின் அடிப்படையில், (படம் 3): Nrf2 இல்லாத நிலையில், ME1, IDH1, G6PD மற்றும் PGD ஆகியவற்றின் வெளிப்பாடு குறைவதால் NADPH அளவுகள் குறைவாக உள்ளன. குறைக்கப்பட்ட குளுதாதயோனின் அளவுகள் அதன் உயிரியக்கவியல் மற்றும் மீளுருவாக்கம் ஆகியவற்றில் பங்குபெறும் நொதிகளின் வெளிப்பாடு குறைவதால் மற்றும் குறைந்த அளவு NADPH காரணமாக ஆக்ஸிஜனேற்றப்பட்டதை குளுதாதயோனின் குறைக்கப்பட்ட வடிவத்திற்கு மாற்றுவதற்குத் தேவைப்படுகிறது. ME1 இன் குறைந்த வெளிப்பாடு மைட்டோகாண்ட்ரியாவுக்குள் நுழையும் பைருவேட்டின் குளத்தை குறைக்கும், கிளைகோலிசிஸ் பைருவேட்டின் முக்கிய ஆதாரமாகிறது. NADH இன் உருவாக்கம் மெதுவாக உள்ளது, இது சிக்கலான I இன் பலவீனமான செயல்பாடு மற்றும் அதிகரித்த மைட்டோகாண்ட்ரியல் ROS உற்பத்திக்கு வழிவகுக்கிறது. FAD ஐ FADH2 ஆக குறைப்பதும் மெதுவாக உள்ளது, குறைந்த பட்சம் குறைந்த செயல்திறன் கொண்ட கொழுப்பு அமில ஆக்சிஜனேற்றம் காரணமாக, FADH2 இலிருந்து UbQ மற்றும் சிக்கலான III க்கு எலக்ட்ரான் ஓட்டத்தை சமரசம் செய்கிறது. UbQH2 என்பது சக்சினேட் டீஹைட்ரோஜினேஸின் [80] செயல்பாட்டாளர் என்பதால், அதன் உருவாக்கத்தை மெதுவாக்குவது சக்சினேட் டீஹைட்ரோஜினேஸின் நொதி செயல்பாட்டைக் குறைக்கலாம். சூப்பர் ஆக்சைடு மற்றும் ஹைட்ரஜன் பெராக்சைட்டின் அதிகரித்த அளவு சிக்கலான II செயல்பாட்டை மேலும் தடுக்கலாம் [81]. கொழுப்பு அமில ஆக்சிஜனேற்றத்தின் குறைந்த செயல்திறன், மைட்டோகாண்ட்ரியல் சுவாசம் மற்றும் ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷனில் ATP உற்பத்திக்கான அடி மூலக்கூறு கிடைப்பதில் குறைவுக்கு பங்களிக்கிறது. இழப்பீட்டு பொறிமுறையாக, கிளைகோலிசிஸ் மேம்படுத்தப்படுகிறது. ATP சின்தேஸ் ??m ஐ பராமரிக்கும் முயற்சியில் ATPase ஆக தலைகீழாக செயல்படுகிறது.
Nrf2 மற்றும் மைட்டோகாண்ட்ரியல் பயோஜெனீசிஸ்
WT உடன் ஒப்பிடும்போது, Nrf2-KO எலிகளின் கல்லீரல்கள் குறைந்த மைட்டோகாண்ட்ரியல் உள்ளடக்கத்தைக் கொண்டுள்ளன (மைட்டோகாண்ட்ரியல் மற்றும் நியூக்ளியர் டிஎன்ஏ விகிதத்தால் தீர்மானிக்கப்படுகிறது); WT மற்றும் Nrf24-KO எலிகள் இரண்டிலும் 2-மணிநேர வேகத்தால் இது மேலும் குறைக்கப்படுகிறது; மாறாக, சாதாரண உணவு நிலைமைகளின் கீழ் WT இலிருந்து வேறுபடவில்லை என்றாலும், அதிக Nrf2 செயல்பாடு கொண்ட எலிகளில் உள்ள மைட்டோகாண்ட்ரியல் உள்ளடக்கம் உண்ணாவிரதத்தால் பாதிக்கப்படுவதில்லை [82]. சுவாரஸ்யமாக, Nrf2 ஆக்டிவேட்டர் (R)-?-லிபோயிக் அமிலம் [83], [84], [85] உடன் கூடுதலாக வழங்குவது 3T3-L1 அடிபோசைட்டுகளில் [86] மைட்டோகாண்ட்ரியல் பயோஜெனீசிஸை ஊக்குவிக்கிறது. மைட்டோகாண்ட்ரியல் பயோஜெனீசிஸில் இரண்டு வகை அணுக்கரு டிரான்ஸ்கிரிப்ஷனல் ரெகுலேட்டர்கள் முக்கிய பங்கு வகிக்கின்றன. முதல் வகுப்பு அணு சுவாசக் காரணிகள்11 மற்றும் 2 போன்ற டிரான்ஸ்கிரிப்ஷன் காரணிகள் ஆகும், இவை ஐந்து சுவாச வளாகங்களின் மரபணு குறியாக்க துணைக்குழுக்கள், மைட்டோகாண்ட்ரியல் மொழிபெயர்ப்பு கூறுகள் மற்றும் மைட்டோகாண்ட்ரியல் மேட்ரிக்ஸில் உள்ளமைக்கப்பட்ட ஹீம் பயோசிந்தெடிக் என்சைம்களின் வெளிப்பாட்டைக் கட்டுப்படுத்துகின்றன [88]. பியாண்டடோசி மற்றும் பலர். [89] நியூக்ளியர் ரெஸ்பிரேட்டரி ஃபேக்டர் 2 இன் Nrf1-சார்ந்த டிரான்ஸ்கிரிப்ஷனல் முறைப்படுத்தல் மைட்டோகாண்ட்ரியல் பயோஜெனீசிஸை ஊக்குவிக்கிறது மற்றும் கார்டியோடாக்ஸிக் ஆந்த்ராசைக்ளின் கீமோதெரபியூடிக் ஏஜென்ட் டாக்ஸோரூபிகின் சைட்டோடாக்ஸிசிட்டிக்கு எதிராக பாதுகாக்கிறது. இதற்கு மாறாக, ஜாங் மற்றும் பலர். [82] Nrf2 இன் மரபணு செயலாக்கமானது முரைன் கல்லீரலில் உள்ள அணுக்கரு சுவாசக் காரணி 1 இன் அடிப்படை mRNA வெளிப்பாட்டை பாதிக்காது என்று கூறியுள்ளனர்.
மைட்டோகாண்ட்ரியல் பயோஜெனீசிஸில் முக்கியமான செயல்பாடுகளைக் கொண்ட அணுக்கரு டிரான்ஸ்கிரிப்ஷனல் ரெகுலேட்டர்களின் இரண்டாம் வகுப்பு டிரான்ஸ்கிரிப்ஷனல் கோஆக்டிவேட்டர்கள், அதாவது பெராக்ஸிசோம் ப்ரோலிஃபெரேட்டர்-ஆக்டிவேட்டட் ரிசெப்டர் ? கோஆக்டிவேட்டர்கள் (PGC)1? மற்றும் 1?, இது டிரான்ஸ்கிரிப்ஷன் காரணிகள், பாசல் டிரான்ஸ்கிரிப்ஷனல் மற்றும் ஆர்என்ஏ-பிளக்கும் இயந்திரங்கள் மற்றும் ஹிஸ்டோனை மாற்றும் என்சைம்கள் [88], [90], [91] ஆகியவற்றுடன் தொடர்பு கொள்கிறது. கோஆக்டிவேட்டர்களின் PGC1 குடும்பத்தின் வெளிப்பாடு பல சுற்றுச்சூழல் சமிக்ஞைகளால் பாதிக்கப்படுகிறது. மனித ஃபைப்ரோபிளாஸ்ட்களை Nrf2 ஆக்டிவேட்டர் சல்ஃபோராபேன் மூலம் சிகிச்சையளிப்பது மைட்டோகாண்ட்ரியல் மாஸ் மற்றும் PGC1 இன் தூண்டலை அதிகரிக்கச் செய்கிறது? மற்றும் PGC1? [92], இந்த ஆய்வில் Nrf2 மீதான சாத்தியமான சார்பு ஆராயப்படவில்லை. இருப்பினும், கீப்2 மரபணு ஹைப்போமார்ஃபிக் நாக் டவுன் (db/db:Keap1flox/?:Nrf1+/+) அல்லது சீர்குலைந்த (db/db:Keap2flox/?:Nrf1?/?) மூலம் Nrf2 செயல்படுத்தப்பட்ட நீரிழிவு எலிகள் குறைந்த கல்லீரல் PGC1 உள்ளதா? கட்டுப்பாட்டு விலங்குகளை விட வெளிப்பாடு நிலைகள் (db/db:Keap1flox/+:Nrf2+/+) [93]. PGC1 க்கான mRNA அளவுகளில் வேறுபாடுகள் இல்லையா? WT அல்லது Nrf2-KO எனப்படும் நீரிழிவு நோயற்ற எலிகளின் கல்லீரலில் காணப்படுகின்றன, அதேசமயம் Nrf2-ஓவர் எக்ஸ்பிரஸ்ஸிங் (Keap1-KD மற்றும் கல்லீரல்-குறிப்பிட்ட Keap1-KO) விலங்குகளில் இந்த அளவுகள் குறைவாக இருக்கும் [82]. குறிப்பிடத்தக்க வகையில், 24-மணிநேர உண்ணாவிரதம் PGC1 இன் அளவை அதிகரிக்குமா? அனைத்து மரபணு வகைகளின் எலிகளின் கல்லீரலில் mRNA உள்ளது, ஆனால் WT அல்லது Nrf2-ஓவர் எக்ஸ்பிரஸ்ஸிங் எலிகளுடன் ஒப்பிடும்போது Nrf2-KO இன் கல்லீரல்களில் அதிகரிப்பு கணிசமாக அதிகமாக உள்ளது. WT உடன் ஒப்பிடும்போது, Nrf2-KO எலிகள் செப்டிக் தொற்று அல்லது தொற்று காரணமாக கடுமையான நுரையீரல் காயத்தை அனுபவிக்கின்றன, அணுக்கரு சுவாச காரணி 1 மற்றும் PGC1 ஆகியவற்றின் டிரான்ஸ்கிரிப்ஷனல் அதிகப்படுத்தலைக் காட்டுகின்றனவா? [94], [95]. ஒன்றாக, இந்த அவதானிப்புகள் அணு சுவாசக் காரணி 2 மற்றும் PGC1 இரண்டின் அளவையும் பராமரிப்பதில் Nrf1 இன் பங்கு என்று கூறுகின்றன. சிக்கலானது மற்றும் மன அழுத்த சூழ்நிலைகளில் மிகவும் முக்கியமானது.
மைட்டோகாண்ட்ரியல் புரதங்களை குறியாக்கம் செய்யும் மரபணுக்களின் வெளிப்பாட்டுடன் கூடுதலாக, மைட்டோகாண்ட்ரியல் உயிரியக்கத்திற்கு நியூக்ளியோடைடுகளின் தொகுப்பு தேவைப்படுகிறது. Nrf2 இன் மரபணு செயலாக்கமானது பென்டோஸ் பாஸ்பேட் பாதை மற்றும் ஃபோலேட் மற்றும் குளுட்டமைனின் வளர்சிதைமாற்றம், குறிப்பாக வேகமாகப் பெருகும் உயிரணுக்களில் (படம் 2) [24] ஆகியவற்றைக் கட்டுப்படுத்துவதன் மூலம் பியூரின் உயிரியக்கத்தை மேம்படுத்துகிறது. மைட்டோகாண்ட்ரியல் செரீன்/திரோயோனைன் புரோட்டீன் கைனேஸ் PTEN-தூண்டப்பட்ட புட்டேட்டிவ் கைனேஸ் 1 (PINK1) க்கான பிறழ்ந்த டிரோசோபிலா குறைபாடுள்ள டிரான்ஸ்கிரிப்டோமின் பகுப்பாய்வு, மைட்டோகாண்ட்ரியல் செயலிழப்பு டிரான்ஸ்கிரிப்ஷனல் மேம்பாட்டிற்கு வழிவகுக்கிறது என்று காட்டுகிறது. PINK96 குறைபாட்டின் நியூரோடாக்ஸிக் விளைவுகளுக்கு எதிரான பாதுகாப்பிற்கான ஒரு பொறிமுறையை பிரதிபலிக்கிறது. Nrf1 பாஸ்போரிபோசைல் பைரோபாஸ்பேட் அமிடோட்ரான்ஸ்ஃபெரேஸின் (PPAT) வெளிப்பாட்டை ஒழுங்குபடுத்துகிறது, இது டி நோவோ பியூரின் நியூக்ளியோடைடு உயிரியக்கவியல் பாதை மற்றும் மைட்டோகாண்ட்ரியல் மெத்திலினெடெட்ராஹைட்ரோஃபோலேட் டீஹைட்ரோஜினேஸ் 2 (MTHFD2) (படம் 2) ஆகியவற்றில் நுழைவதை ஊக்குவிக்கிறது. பிந்தையது டீஹைட்ரோஜினேஸ் மற்றும் சைக்ளோஹைட்ரோலேஸ் செயல்பாடுகளுடன் கூடிய இருசெயல்பாட்டு நொதியாகும், இது வேகமாக வளரும் உயிரணுக்களில் பியூரின் உயிரியக்கத்திற்கு ஒரு கார்பன் அலகுகளின் ஆதாரமாக கிளைசின் மற்றும் ஃபார்மேட் இரண்டையும் வழங்குவதில் முக்கியமானது [2]. எனவே Nrf97 செயல்படுத்தல் பாதுகாப்பாக இருக்கலாம் மற்றும் PINK2 குறைபாட்டில் மைட்டோகாண்ட்ரியல் செயலிழப்பை மாற்றியமைக்கலாம். உண்மையில், சல்ஃபோராபேன் அல்லது ட்ரைடர்பெனாய்டு RTA-1 மூலம் Nrf2 இன் மருந்தியல் செயலாக்கம் ??m மீட்டமைக்கிறது மற்றும் டோபமைன் நச்சுத்தன்மைக்கு எதிராக PINK408-குறைபாடுள்ள செல்களைப் பாதுகாக்கிறது [1]. அடிப்படை வழிமுறைகள் சிக்கலானதாகத் தோன்றினாலும், ஒன்றாக, இந்த கண்டுபிடிப்புகள் Nrf98 செயல்பாடு மைட்டோகாண்ட்ரியல் பயோஜெனீசிஸை முக்கியமான டிரான்ஸ்கிரிப்ஷன் காரணிகள் மற்றும் கோஆக்டிவேட்டர்களின் வெளிப்பாடு நிலைகளில் செல்வாக்கு செலுத்துவதன் மூலம் மற்றும் நியூக்ளியோடைடு உயிரியக்கத்தை மேம்படுத்துவதன் மூலம் பாதிக்கலாம் என்பதைக் குறிக்கிறது.
Nrf2 மற்றும் மைட்டோகாண்ட்ரியல் ஒருமைப்பாடு
நேரடி சான்றுகள் எப்போதும் கிடைக்காது என்றாலும், மைட்டோகாண்ட்ரியல் ஒருமைப்பாட்டிற்கு, குறிப்பாக ஆக்ஸிஜனேற்ற அழுத்தத்தின் கீழ் Nrf2 முக்கியமானது என்பதற்கான வலுவான அறிகுறிகள் உள்ளன. Nrf2 ஆக்டிவேட்டர் சல்ஃபோராபேன் ஒரு டோஸ் கொடுக்கப்பட்ட எலிகளின் மூளை மற்றும் கல்லீரலில் இருந்து தனிமைப்படுத்தப்பட்ட மைட்டோகாண்ட்ரியா, டெர்ட்-பியூட்டில்ஹைட்ரோபெராக்சைடு [99] [100], [1500] என்ற ஆக்ஸிஜனேற்றத்தால் ஏற்படும் மைட்டோகாண்ட்ரியல் பெர்மபிலிட்டி டிரான்சிஷன் போரை (mPTP) திறப்பதை எதிர்க்கும். mPTP, மைட்டோகாண்ட்ரியல் உள் சவ்வு 0 Da வரை நிறை கொண்ட மூலக்கூறுகளுக்கு ஊடுருவ அனுமதிக்கும் ஒரு சிக்கலானது, F1F101-ATP சின்தேஸின் டைமர்களில் இருந்து உருவாகிறது என்று சமீபத்தில் கண்டறியப்பட்டது [1]. mPTP திறப்புக்கான சல்ஃபோராபேன்-மத்தியஸ்த எதிர்ப்பு, அதிகரித்த ஆக்ஸிஜனேற்ற பாதுகாப்புகளுடன் தொடர்புபடுத்துகிறது, மேலும் மைட்டோகாண்ட்ரியல் GSH, குளுதாதயோன் பெராக்சிடேஸ் 3, மாலிக் என்சைம் 2 மற்றும் தியோரெடாக்சின் 100 ஆகியவற்றின் அளவுகள் அனைத்தும் மைட்டோகாண்ட்ரியல் பின்னங்களில் கட்டுப்படுத்தப்படுகின்றன [sultreaphane-XNUMX]
மைட்டோகாண்ட்ரியல் புரதச் சேதம் மற்றும் எலெக்ட்ரோஃபிலிக் லிப்பிட் பெராக்ஸைடேஷன் தயாரிப்பு 4-ஹைட்ராக்ஸி-2-நோனெனல் காரணமாக ஏற்படும் சுவாசக் குறைபாடு ஆகியவை சல்ஃபோராபேன்-சிகிச்சையளிக்கப்பட்ட எலிகளின் பெருமூளைப் புறணியில் இருந்து தனிமைப்படுத்தப்பட்ட மைட்டோகாண்ட்ரியாவில் குறைக்கப்படுகின்றன [102]. எலி சிறுநீரக எபிடெலியல் செல்கள் மற்றும் சிறுநீரகத்தில், சல்ஃபோராபேன் சிஸ்ப்ளேட்டின்- மற்றும் ஜென்டாமைசின்-தூண்டப்பட்ட நச்சுத்தன்மை மற்றும் ??மீ[103], [104] இழப்பிற்கு எதிராக பாதுகாக்கிறது. ஆக்சிடென்ட்கள் (சூப்பராக்சைடு, ஹைட்ரஜன் பெராக்சைடு, பெராக்ஸைனிட்ரைட்) மற்றும் எலக்ட்ரோஃபில்ஸ் (4-ஹைட்ராக்ஸி-2-நோனெனல் மற்றும் அக்ரோலின்) ஆகியவற்றிற்கு எதிரான பாதுகாப்பு மற்றும் மைட்டோகாண்ட்ரியல் ஆக்ஸிஜனேற்ற பாதுகாப்பு அதிகரிப்பு ஆகியவை எலி பெருநாடியின் மென்மையான தசை செல்களை சல்ஃபோராபேன் [105] மூலம் சிகிச்சையளிப்பதன் மூலம் கவனிக்கப்பட்டது. ]. கான்ட்ராஸ்ட்-தூண்டப்பட்ட கடுமையான சிறுநீரக காயத்தின் மாதிரியில், ஜிஎஸ்கே 2 இன் தடுப்பின் விளைவாக Nrf3 ஐ செயல்படுத்துவதன் மூலம், mPTP மற்றும் மைட்டோகாண்ட்ரியல் வீக்கத்தைத் திறப்பதைத் தடுப்பது உள்ளிட்ட பாதுகாப்பு விளைவுகளை மூட்டு இஸ்கிமிக் முன்நிபந்தனை சமீபத்தில் காட்டப்பட்டது? [106].
மைட்டோபாகி, செயலிழந்த மைட்டோகாண்ட்ரியாவை ஆட்டோபாகோசோம்களால் தேர்ந்தெடுக்கப்பட்டு, லைசோசோம்களுக்கு அனுப்பப்பட்டு, உயிரணுவால் சிதைக்கப்பட்டு மறுசுழற்சி செய்யப்படும், இது மைட்டோகாண்ட்ரியல் ஹோமியோஸ்டாசிஸ் [107], [108] இன்றியமையாதது. Nrf2 மற்றும் mitophagy இடையே எந்த காரணமான தொடர்பும் நிறுவப்படவில்லை என்றாலும், mitochondrial தரக் கட்டுப்பாட்டில் டிரான்ஸ்கிரிப்ஷன் காரணி முக்கியமானதாக இருக்கலாம் என்பதற்கான சான்றுகள் உள்ளன. ஆக்ஸிஜனேற்ற அழுத்தத்தின் நிலைமைகளின் கீழ் இது குறிப்பாக முக்கியத்துவம் வாய்ந்ததாக இருக்கலாம். இவ்வாறு, செப்சிஸின் மாதிரியில், தன்னியக்க மார்க்கர் MAP1 லைட் செயின் 3-II (LC3-II) மற்றும் கார்கோ புரதம் p62 ஆகியவற்றின் அளவுகள் 24 h போஸ்ட் இன்ஃபெக்ஷனில் WT எலிகளுடன் ஒப்பிடும்போது Nrf2-KO இல் அடக்கப்படுகின்றன [109] . மைட்டோபாகியின் ஒரு சிறிய-மூலக்கூறு தூண்டி (p62-மத்தியஸ்த மைட்டோபாகி தூண்டி, PMI என்று அழைக்கப்படுகிறது) சமீபத்தில் கண்டுபிடிக்கப்பட்டது; இந்த 1,4-டிஃபெனைல்-1,2,3-ட்ரையசோல் கலவை முதலில் Nrf2 ஆக்டிவேட்டராக வடிவமைக்கப்பட்டது, இது கீப்1 [110] உடன் டிரான்ஸ்கிரிப்ஷன் காரணியின் தொடர்புகளை சீர்குலைக்கிறது. Nrf2 மரபணு ரீதியாக கட்டுப்படுத்தப்பட்ட செல்களைப் போலவே (Keap1-KD அல்லது Keap1-KO), PMI க்கு வெளிப்படும் செல்கள் அதிக ஓய்வு ??m. முக்கியமாக, WT செல்களின் PMI சிகிச்சைக்குப் பிறகு காணப்படும் மைட்டோகாண்ட்ரியல் LC3 உள்ளூர்மயமாக்கலின் அதிகரிப்பு Nrf2-KO கலங்களில் ஏற்படாது, இது Nrf2 இன் ஈடுபாட்டைக் குறிக்கிறது.
கடைசியாக, கல்லீரல் பிரிவுகளின் அல்ட்ராஸ்ட்ரக்சரல் பகுப்பாய்வு, 2 வாரங்களுக்கு அதிக கொழுப்புள்ள உணவைக் கொடுத்த எலிகளுக்கு Nrf24-KO இன் ஹெபடோசைட்டுகளில் குறைந்த கிரிஸ்டா மற்றும் சீர்குலைந்த சவ்வுகளுடன் வீங்கிய மைட்டோகாண்ட்ரியா இருப்பதை வெளிப்படுத்தியுள்ளது, ஆனால் WT அல்ல; குறிப்பாக, இந்த கல்லீரல்கள் ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் அழற்சியின் தெளிவான ஆதாரங்களைக் காட்டுகின்றன [68]. ஆக்ஸிஜனேற்ற மற்றும் அழற்சி அழுத்தத்தின் கீழ் மைட்டோகாண்ட்ரியல் ஒருமைப்பாட்டை பராமரிப்பதில் Nrf2 ஒரு முக்கிய பங்கைக் கொண்டுள்ளது என்று முடிவு செய்யலாம்.
புற்றுநோய், இறப்பு, முதுமை, மூளை மற்றும் நடத்தை, இதய நோய் மற்றும் பலவற்றில் சல்போராபேன் மற்றும் அதன் விளைவுகள்
ஐசோதியோசயனேட்டுகள் உங்கள் உணவில் நீங்கள் பெறக்கூடிய மிக முக்கியமான தாவர கலவைகள் ஆகும். இந்த வீடியோவில் நான் அவர்களுக்காக இதுவரை செய்யப்படாத மிக விரிவான வழக்கை உருவாக்குகிறேன். குறுகிய கவனம்? கீழே உள்ள நேரப் புள்ளிகளில் ஒன்றைக் கிளிக் செய்வதன் மூலம் உங்களுக்குப் பிடித்த தலைப்புக்குச் செல்லவும். முழு காலவரிசை கீழே.
முக்கிய பிரிவுகள்:
00:01:14 - புற்றுநோய் மற்றும் இறப்பு
00:19:04 - முதுமை
00:26:30 - மூளை மற்றும் நடத்தை
00:38:06 - இறுதி மறுபரிசீலனை
00:40:27 - டோஸ்
முழு காலவரிசை:
00:00:34 – வீடியோவின் முக்கிய மையமான சல்போராபேன் அறிமுகம்.
00:01:14 - க்ரூசிஃபெரஸ் காய்கறி நுகர்வு மற்றும் அனைத்து காரணங்களின் இறப்பு குறைப்பு.
00:02:12 - புரோஸ்டேட் புற்றுநோய் ஆபத்து.
00:02:23 - சிறுநீர்ப்பை புற்றுநோய் ஆபத்து.
00:02:34 - புகைப்பிடிப்பவர்களுக்கு நுரையீரல் புற்றுநோய் ஆபத்து.
00:02:48 - மார்பக புற்றுநோய் ஆபத்து.
00:03:13 - அனுமானம்: உங்களுக்கு ஏற்கனவே புற்றுநோய் இருந்தால் என்ன செய்வது? (இடையிடல்)
00:03:35 – புற்றுநோய் மற்றும் இறப்பு தொடர்பான தரவுகளை இயக்கும் நம்பத்தகுந்த வழிமுறை.
00:04:38 - சல்போராபேன் மற்றும் புற்றுநோய்.
00:05:32 - எலிகளில் சிறுநீர்ப்பை கட்டி வளர்ச்சியில் ப்ரோக்கோலி முளை சாறு வலுவான விளைவைக் காட்டும் விலங்கு சான்றுகள்.
00:06:06 - புரோஸ்டேட் புற்றுநோயாளிகளில் சல்ஃபோராபேன் நேரடியாக நிரப்புவதன் விளைவு.
00:07:09 - உண்மையான மார்பக திசுக்களில் ஐசோதியோசயனேட் வளர்சிதை மாற்றங்களின் உயிர் குவிப்பு.
00:08:32 - மார்பக புற்றுநோய் ஸ்டெம் செல்களைத் தடுப்பது.
00:08:53 - வரலாற்றுப் பாடம்: பண்டைய ரோமில் கூட பிராசிகாக்கள் ஆரோக்கிய பண்புகளைக் கொண்டிருப்பதாக நிறுவப்பட்டது.
00:09:16 – சல்போராபேன் புற்றுநோய் வெளியேற்றத்தை மேம்படுத்தும் திறன் (பென்சீன், அக்ரோலின்).
00:09:51 - ஆன்டிஆக்ஸிடன்ட் மறுமொழி கூறுகள் வழியாக மரபணு மாற்றமாக NRF2.
00:10:10 – NRF2 ஆக்டிவேஷன் குளுதாதயோன்-எஸ்-கான்ஜுகேட்ஸ் வழியாக புற்றுநோயை வெளியேற்றுவதை எவ்வாறு மேம்படுத்துகிறது.
00:10:34 - பிரஸ்ஸல்ஸ் முளைகள் குளுதாதயோன்-எஸ்-டிரான்ஸ்ஃபெரேஸை அதிகரிக்கின்றன மற்றும் டிஎன்ஏ பாதிப்பைக் குறைக்கின்றன.
00:11:20 - ப்ரோக்கோலி முளை பானம் பென்சீன் வெளியேற்றத்தை 61% அதிகரிக்கிறது.
00:13:31 - ப்ரோக்கோலி ஸ்ப்ரூட் ஹோமோஜெனேட் மேல் சுவாசப்பாதையில் ஆக்ஸிஜனேற்ற நொதிகளை அதிகரிக்கிறது.
00:15:45 - சிலுவை காய்கறி நுகர்வு மற்றும் இதய நோய் இறப்பு.
00:16:55 - ப்ரோக்கோலி ஸ்ப்ரூட் பவுடர் இரத்த கொழுப்பு மற்றும் ஒட்டுமொத்த இதய நோய் அபாயத்தை மேம்படுத்துகிறது வகை 2 நீரிழிவு நோயாளிகள்.
00:19:04 - வயதான பிரிவின் ஆரம்பம்.
00:19:21 - சல்போராபேன்-செறிவூட்டப்பட்ட உணவு வண்டுகளின் ஆயுட்காலம் 15 முதல் 30% வரை அதிகரிக்கிறது (சில நிபந்தனைகளில்).
00:20:34 - நீண்ட ஆயுளுக்கு குறைந்த வீக்கத்தின் முக்கியத்துவம்.
00:22:05 - குரூசிஃபெரஸ் காய்கறிகள் மற்றும் ப்ரோக்கோலி ஸ்ப்ரூட் பவுடர் மனிதர்களில் பல்வேறு வகையான அழற்சி குறிப்பான்களைக் குறைப்பதாகத் தெரிகிறது.
00:23:40 – மிட்-வீடியோ ரீகேப்: புற்றுநோய், வயதான பிரிவுகள்
00:24:14 - முதுமையில் சல்போராபேன் தகவமைப்பு நோயெதிர்ப்புச் செயல்பாட்டை மேம்படுத்தலாம் என்று சுட்டி ஆய்வுகள் தெரிவிக்கின்றன.
00:25:18 – சல்ஃபோராபேன், வழுக்கை மவுஸ் மாதிரியில் முடி வளர்ச்சியை மேம்படுத்தியது. படம் 00:26:10.
00:28:17 - மனச்சோர்வு விவாதத்தின் தொடக்கம் (நம்பத்தகுந்த வழிமுறை மற்றும் ஆய்வுகள்).
00:31:21 – அழுத்தத்தால் தூண்டப்பட்ட மனச்சோர்வின் 10 வெவ்வேறு மாதிரிகளைப் பயன்படுத்தி மவுஸ் ஆய்வு, ஃப்ளூக்ஸெடின் (ப்ரோசாக்) போலவே சல்ஃபோராபேன் பயனுள்ளதாக இருக்கும்.
00:32:00 – எலிகளில் குளுக்கோராபனின் நேரடியாக உட்கொள்வது சமூக தோல்வி மன அழுத்த மாதிரியிலிருந்து மனச்சோர்வைத் தடுப்பதில் இதேபோல் பயனுள்ளதாக இருக்கும் என்று ஆய்வு காட்டுகிறது.
00:33:01 - நியூரோடிஜெனரேஷன் பிரிவின் ஆரம்பம்.
00:33:30 – சல்போராபேன் மற்றும் அல்சைமர் நோய்.
00:33:44 – சல்போராபேன் மற்றும் பார்கின்சன் நோய்.
00:33:51 – சல்போராபேன் மற்றும் ஹங்டிங்டன் நோய்.
00:34:13 - சல்ஃபோராபேன் வெப்ப அதிர்ச்சி புரதங்களை அதிகரிக்கிறது.
00:34:43 - அதிர்ச்சிகரமான மூளை காயம் பிரிவின் ஆரம்பம்.
00:35:55 - சல்ஃபோராபேன் மற்றும் நியூரானல் பிளாஸ்டிசிட்டி.
00:36:32 – எலிகளில் டைப் II நீரிழிவு மாதிரியில் சல்போராபேன் கற்றலை மேம்படுத்துகிறது.
00:37:19 - சல்போராபேன் மற்றும் டுச்சேன் தசைநார் சிதைவு.
00:37:44 - தசை செயற்கைக்கோள் செல்களில் Myostatin தடுப்பு (விட்ரோவில்).
00:38:06 – லேட்-வீடியோ மறுபரிசீலனை: இறப்பு மற்றும் புற்றுநோய், டிஎன்ஏ சேதம், ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் வீக்கம், பென்சீன் வெளியேற்றம், இருதய நோய், வகை II நீரிழிவு, மூளையில் விளைவுகள் (மன அழுத்தம், மன இறுக்கம், ஸ்கிசோஃப்ரினியா, நியூரோடிஜெனரேஷன்), NRF2 பாதை.
00:40:27 - ப்ரோக்கோலி முளைகள் அல்லது சல்ஃபோராபேன் அளவைக் கண்டறிவது பற்றிய எண்ணங்கள்.
00:41:01 - வீட்டில் முளைப்பது பற்றிய நிகழ்வுகள்.
00:43:14 - சமையல் வெப்பநிலை மற்றும் சல்ஃபோராபேன் செயல்பாடு.
00:43:45 - குளுகோராபனினில் இருந்து சல்போராபேன் குடல் பாக்டீரியாவை மாற்றுகிறது.
00:44:24 - காய்கறிகளிலிருந்து செயலில் உள்ள மைரோசினேஸுடன் இணைந்தால் சப்ளிமெண்ட்ஸ் சிறப்பாகச் செயல்படும்.
00:44:56 - சமையல் நுட்பங்கள் மற்றும் சிலுவை காய்கறிகள்.
00:46:06 - ஐசோதியோசயனேட்டுகள் கோயிட்ரோஜன்களாக.
Nrf2 என்பது டிரான்ஸ்கிரிப்ஷன் காரணியாகும், இது மனித உடலின் செல்லுலார் ஆக்ஸிஜனேற்ற பாதுகாப்பு அமைப்பில் முக்கிய பங்கு வகிக்கிறது. ஆக்ஸிஜனேற்ற பதிலளிக்கக்கூடிய உறுப்பு, அல்லது ARE, மரபணுக்களின் ஒழுங்குமுறை பொறிமுறையாகும். Nrf2, அல்லது NF-E2-தொடர்புடைய காரணி 2, பல வகையான செல்கள் முழுவதும் பலவிதமான ARE-உந்துதல் மரபணுக்களை ஒழுங்குபடுத்துகிறது என்பதை பல ஆராய்ச்சி ஆய்வுகள் நிரூபித்துள்ளன. Nrf2 செல்லுலார் பாதுகாப்பு மற்றும் புற்றுநோய் எதிர்ப்புத் தன்மையில் முக்கியப் பங்கு வகிப்பதாகக் கண்டறியப்பட்டது, இது ஆக்ஸிஜனேற்ற அழுத்தத்தால் ஏற்படக்கூடிய நரம்பியக்கடத்தல் நோய்கள் மற்றும் புற்றுநோய்களை நிர்வகிப்பதில் Nrf2 ஒரு சிறந்த சிகிச்சையாக இருக்கலாம் என்பதை நிரூபிக்கிறது. டாக்டர். அலெக்ஸ் ஜிமினெஸ் DC, CCST இன்சைட்
இறுதியான குறிப்புகள்
பல கேள்விகள் இன்னும் திறந்தே இருந்தாலும், மைட்டோகாண்ட்ரியல் ஹோமியோஸ்டாஸிஸ் மற்றும் கட்டமைப்பு ஒருமைப்பாட்டைப் பராமரிப்பதில் Nrf2 ஒரு முக்கிய பங்கு வகிக்கிறது என்பதை தற்போதுள்ள சோதனை ஆதாரங்கள் தெளிவாகக் குறிப்பிடுகின்றன. Nrf2-மத்தியஸ்த சைட்டோபுரோடெக்டிவ் பதில்களைக் கட்டுப்படுத்தும் திறன் உயிரணு மற்றும் உயிரினத்தின் ஒட்டுமொத்த ஆரோக்கியம் மற்றும் உயிர்வாழ்வை பாதிக்கும் போது ஆக்ஸிஜனேற்ற, எலக்ட்ரோஃபிலிக் மற்றும் அழற்சி அழுத்தத்தின் நிலைமைகளின் கீழ் இந்த பாத்திரம் குறிப்பாக முக்கியமானதாகிறது. மைட்டோகாண்ட்ரியல் செயல்பாட்டில் Nrf2 இன் பங்கு, இந்த டிரான்ஸ்கிரிப்ஷன் காரணியால் திட்டமிடப்பட்ட பரந்த சைட்டோபுரோடெக்டிவ் வழிமுறைகளின் மற்றொரு அடுக்கைக் குறிக்கிறது. பல மனித நோயியல் நிலைமைகள் ஆக்ஸிஜனேற்ற அழுத்தம், வீக்கம் மற்றும் மைட்டோகாண்ட்ரியல் செயலிழப்பு ஆகியவை அவற்றின் நோய்க்கிருமிகளின் முக்கிய கூறுகளாக இருப்பதால், Nrf2 இன் மருந்தியல் செயல்படுத்தல் நோய் தடுப்பு மற்றும் சிகிச்சைக்கான உறுதிமொழியைக் கொண்டுள்ளது. Nrf2 மைட்டோகாண்ட்ரியல் செயல்பாட்டைப் பாதிக்கும் துல்லியமான வழிமுறைகளைப் பற்றிய விரிவான புரிதல் எதிர்கால மருத்துவ பரிசோதனைகளின் பகுத்தறிவு வடிவமைப்பிற்கு அவசியமானது மற்றும் சிகிச்சை செயல்திறனைக் கண்காணிக்க புதிய பயோமார்க்ஸர்களை வழங்கலாம்.
மேலே உள்ள கட்டுரையின் நோக்கம், மைட்டோகாண்ட்ரியல் செயல்பாட்டில் Nrf2 இன் வளர்ந்து வரும் பங்கைப் பற்றி விவாதிப்பதும், நிரூபிப்பதும் ஆகும். Nrf2, அல்லது அணுக்கரு காரணி எரித்ராய்டு 2 தொடர்பான காரணி, ஆக்ஸிஜனேற்ற அழுத்தத்திற்கு பங்களிக்கும், செல்லுலார் செயல்பாட்டை பாதிக்கும் மற்றும் நச்சுத்தன்மை, நாட்பட்ட நோய் மற்றும் புற்றுநோயின் வளர்ச்சிக்கு வழிவகுக்கும் ஆக்ஸிஜனேற்றங்களுக்கு செல்லுலார் எதிர்ப்பின் வளர்ந்து வரும் சீராக்கி. மனித உடலில் ஆக்ஸிஜனேற்றிகளின் உற்பத்தி செல் பிரிவு, வீக்கம், நோயெதிர்ப்பு செயல்பாடு, தன்னியக்க மற்றும் மன அழுத்த பதில் உள்ளிட்ட பல்வேறு நோக்கங்களுக்கு சேவை செய்ய முடியும் என்றாலும், உடல்நலப் பிரச்சினைகளைத் தடுக்க அவற்றின் அதிகப்படியான உற்பத்தியைக் கட்டுப்படுத்துவது அவசியம். எங்கள் தகவலின் நோக்கம் உடலியக்க மற்றும் முதுகெலும்பு சுகாதார பிரச்சினைகள் மட்டுமே. விஷயத்தைப் பற்றி விவாதிக்க, தயவு செய்து டாக்டர் ஜிமினெஸைக் கேட்கவும் அல்லது எங்களை இங்கே தொடர்பு கொள்ளவும்915-850-0900.
முதுகு வலிஉலகளவில் இயலாமை மற்றும் வேலை நாட்களைத் தவறவிடுவதற்கு மிகவும் பொதுவான காரணங்களில் ஒன்றாகும். முதுகுவலியானது மருத்துவர் அலுவலகத்திற்குச் செல்வதற்கான இரண்டாவது பொதுவான காரணமாகும், இது மேல் சுவாச நோய்த்தொற்றுகளால் மட்டுமே அதிகமாக உள்ளது. ஏறக்குறைய 80 சதவீத மக்கள் தங்கள் வாழ்நாள் முழுவதும் ஒருமுறையாவது முதுகுவலியை அனுபவிப்பார்கள். முதுகெலும்பு என்பது மற்ற மென்மையான திசுக்களில் எலும்புகள், மூட்டுகள், தசைநார்கள் மற்றும் தசைகள் ஆகியவற்றால் ஆன ஒரு சிக்கலான அமைப்பாகும். இதன் காரணமாக, காயங்கள் மற்றும்/அல்லது மோசமான நிலைமைகள் போன்றவைஹெர்னியேட்டட் டிஸ்க்குகள், இறுதியில் முதுகுவலியின் அறிகுறிகளுக்கு வழிவகுக்கும். விளையாட்டு காயங்கள் அல்லது வாகன விபத்து காயங்கள் பெரும்பாலும் முதுகுவலிக்கு அடிக்கடி காரணமாகும், இருப்பினும், சில நேரங்களில் எளிமையான இயக்கங்கள் வலிமிகுந்த விளைவுகளை ஏற்படுத்தும். அதிர்ஷ்டவசமாக, உடலியக்க சிகிச்சை போன்ற மாற்று சிகிச்சை விருப்பங்கள், முதுகெலும்பு சரிசெய்தல் மற்றும் கைமுறை கையாளுதல்கள் மூலம் முதுகுவலியை எளிதாக்க உதவும், இறுதியில் வலி நிவாரணத்தை மேம்படுத்துகிறது. �
nrf2 அல்சைமர் நோய் போன்ற ஆக்ஸிஜனேற்ற அழுத்தத்தின் அதிகரித்த அளவுகளுடன் தொடர்புடைய உடல்நலப் பிரச்சினைகளின் விளைவுகளிலிருந்து மனித உடலைப் பாதுகாக்கும் ஆக்ஸிஜனேற்ற மற்றும் நச்சுத்தன்மையற்ற என்சைம்கள் மற்றும் மரபணுக்களின் குழுவை செயல்படுத்துவதை ஆதரிக்கிறது. Nrf2 பாதையை செயல்படுத்த பல்வேறு இயற்கை பொருட்கள் நிரூபிக்கப்பட்டுள்ளன, இது நரம்பியக்கடத்தல் நோய்களின் அறிகுறிகளை நிர்வகிக்க உதவும். கீழேயுள்ள கட்டுரையின் நோக்கம் நாள்பட்ட அழற்சியால் ஏற்படும் Nrf2 இன் முக்கிய பங்கைப் பற்றி விவாதிப்பதாகும்.
சுருக்கம்
பல நாள்பட்ட நோய்கள் மற்றும் சிக்கல்களின் பொதுவான அம்சம் வீக்கம் ஆகும், அதே நேரத்தில் புற்றுநோயை உண்டாக்குவதில் முக்கிய பங்கு வகிக்கிறது. அழற்சி செல்களை ஆட்சேர்ப்பு செய்வதன் மூலம் மற்றும் ஆக்ஸிஜனேற்ற மறுமொழி உறுப்பு (ARE) மூலம் மரபணு வெளிப்பாட்டைக் கட்டுப்படுத்துவதன் மூலம் அழற்சி எதிர்ப்பு செயல்முறைக்கு Nrf2 பங்களிக்கிறது என்பதை பல ஆய்வுகள் நிரூபித்துள்ளன. Keap1 (Kelch-போன்ற ECH-தொடர்புடைய புரதம்)/Nrf2 (NF-E2 p45-தொடர்புடைய காரணி 2)/ARE சிக்னலிங் பாதை முக்கியமாக அழற்சி எதிர்ப்பு மரபணு வெளிப்பாட்டைக் கட்டுப்படுத்துகிறது மற்றும் வீக்கத்தின் முன்னேற்றத்தைத் தடுக்கிறது. எனவே, புதிய Nrf2-சார்ந்த அழற்சி எதிர்ப்பு பைட்டோ கெமிக்கல்களின் அடையாளம் மருந்து கண்டுபிடிப்பில் ஒரு முக்கிய புள்ளியாக மாறியுள்ளது. இந்த மதிப்பாய்வில், Keap1/Nrf2/ARE சிக்னல் பாதையின் உறுப்பினர்கள் மற்றும் அதன் கீழ்நிலை மரபணுக்கள், அழற்சி நோய்களின் விலங்கு மாதிரிகளில் இந்த பாதையின் விளைவுகள் மற்றும் NF-?B பாதையுடன் க்ரோஸ்டாக் பற்றி விவாதிக்கிறோம். கூடுதலாக, NRf3 ஆல் NLRP2 அழற்சியைக் கட்டுப்படுத்துவது பற்றியும் விவாதிக்கிறோம். இது தவிர, Nrf2/ARE சிக்னலிங் பாதைக்கு மத்தியஸ்தம் செய்யும் அழற்சி எதிர்ப்பு பைட்டோ கெமிக்கல்கள் மற்றும் பிறவற்றின் வளர்ச்சியின் தற்போதைய சூழ்நிலையை நாங்கள் சுருக்கமாகக் கூறுகிறோம்.
முக்கிய வார்த்தைகள்: Nrf2, Keap1, ARE, அழற்சி, ஆக்ஸிஜனேற்ற அழுத்தம், பைட்டோகெமிக்கல்
அழற்சி என்பது ஒரு சிக்கலான செயல்முறையாகும், இது நோய்க்கிருமிகள், சேதம் அல்லது எரிச்சல் போன்ற தீங்கு விளைவிக்கும் தூண்டுதல்களால் திசுக்கள் பாதிக்கப்படும் போது அல்லது காயமடைகிறது. நோயெதிர்ப்பு செல்கள், இரத்த நாளங்கள் மற்றும் மூலக்கூறு மத்தியஸ்தர்கள் இந்த பாதுகாப்பு பதிலில் ஈடுபட்டுள்ளனர் [1]. வீக்கம் என்பது உடல், வேதியியல், உயிரியல் மற்றும் உளவியல் காரணிகளால் தூண்டப்படும் பல்வேறு நோய் நிலைகளுடன் தொடர்புடைய ஒரு நோயியல் நிகழ்வு ஆகும். வீக்கத்தின் நோக்கம், செல்லுலார் சேதத்திற்கான காரணங்களைக் கட்டுப்படுத்துவது மற்றும் அகற்றுவது, அழிக்கும் மற்றும்/அல்லது நக்ரோடிக் செல்கள் மற்றும் திசுக்களை உறிஞ்சி, திசு சரிசெய்தலைத் தொடங்குவது. அழற்சியின் இரண்டு வெவ்வேறு வடிவங்கள் வேறுபடுகின்றன: கடுமையான மற்றும் நாள்பட்ட. கடுமையான அழற்சியானது தன்னைத்தானே கட்டுப்படுத்துகிறது மற்றும் ஹோஸ்டுக்கு நன்மை பயக்கும், ஆனால் நீண்ட நாள்பட்ட அழற்சியானது பல நாட்பட்ட நோய்கள் மற்றும் சிக்கல்களின் பொதுவான அம்சமாகும். மோனோசைட்டுகள், மேக்ரோபேஜ்கள், லிம்போசைட்டுகள் மற்றும் பிளாஸ்மா செல்கள் போன்ற பல மோனோநியூக்ளியர் நோயெதிர்ப்பு உயிரணுக்களின் நேரடி ஊடுருவல், அத்துடன் அழற்சி சைட்டோகைன்களின் உற்பத்தி ஆகியவை நாள்பட்ட அழற்சிக்கு வழிவகுக்கும். புற்றுநோயை உண்டாக்குவதில் நாள்பட்ட அழற்சி முக்கிய பங்கு வகிக்கிறது [2]. பொதுவாக, சார்பு மற்றும் அழற்சி எதிர்ப்பு சமிக்ஞை பாதைகள் இரண்டும் சாதாரண அழற்சி செயல்பாட்டில் தொடர்பு கொள்கின்றன.
நோயியல் அழற்சி செயல்பாட்டில், மாஸ்ட் செல்கள், மோனோசைட்டுகள், மேக்ரோபேஜ்கள், லிம்போசைட்டுகள் மற்றும் பிற நோயெதிர்ப்பு செல்கள் முதலில் செயல்படுத்தப்படுகின்றன. பின்னர் உயிரணுக்கள் காயம் ஏற்பட்ட இடத்திற்கு ஆட்சேர்ப்பு செய்யப்படுகின்றன, இதன் விளைவாக டிஎன்ஏ உள்ளிட்ட பெரிய மூலக்கூறுகளை சேதப்படுத்தும் எதிர்வினை ஆக்ஸிஜன் இனங்கள் (ROS) உருவாகின்றன. அதே நேரத்தில், இந்த அழற்சி செல்கள் சைட்டோகைன்கள், கெமோக்கின்கள் மற்றும் புரோஸ்டாக்லாண்டின்கள் போன்ற பெரிய அளவிலான அழற்சி மத்தியஸ்தர்களை உருவாக்குகின்றன. இந்த மத்தியஸ்தர்கள் மேக்ரோபேஜ்களை வீக்கத்தின் உள்ளூர்மயமாக்கப்பட்ட தளங்களுக்கு மேலும் ஆட்சேர்ப்பு செய்கிறார்கள் மற்றும் பல சமிக்ஞை கடத்தும் அடுக்குகள் மற்றும் அழற்சியுடன் தொடர்புடைய டிரான்ஸ்கிரிப்ஷன் காரணிகளை நேரடியாக செயல்படுத்துகிறார்கள். NF-?B (அணு காரணி கப்பா B), MAPK (மைட்டோஜென்-செயல்படுத்தப்பட்ட புரத கைனேஸ்), மற்றும் JAK (ஜானஸ் கைனேஸ்) -STAT (சிக்னல் டிரான்ஸ்டியூசர்கள் மற்றும் டிரான்ஸ்கிரிப்ஷனின் ஆக்டிவேட்டர்கள்) சமிக்ஞை பாதைகள் அழற்சியின் கிளாசிக்கல் பாதையின் வளர்ச்சியில் ஈடுபட்டுள்ளன. [3], [4], [5]. டிரான்ஸ்கிரிப்ஷன் காரணி Nrf2 (NF-E2 p45-தொடர்புடைய காரணி 2) NADPH, NAD(P)H குயினோன் ஆக்சிடோரேடக்டேஸ் 1, குளுதாதயோன் பெராக்சிடேஸ், ஃபெரிடின், ஹீம் ஆக்சிஜனேஸ்-1 உள்ளிட்ட இரண்டாம் நிலை நச்சு நீக்கும் நொதிகளின் வெளிப்பாட்டைக் கட்டுப்படுத்துகிறது என்பதை முந்தைய ஆய்வுகள் வெளிப்படுத்தியுள்ளன. -1), மற்றும் ஆன்டி-ஆக்ஸிடன்ட் ஜீன்கள், பல்வேறு காயங்களிலிருந்து செல்களைப் பாதுகாக்கும் அழற்சி எதிர்ப்பு விளைவுகளால், இதனால் நோயின் போக்கை பாதிக்கிறது [6], [7], [8].
இந்த குறிப்பிடத்தக்க கண்டுபிடிப்புகளைக் கருத்தில் கொண்டு, சமிக்ஞை செய்யும் பாதைகள் மூலம் அழற்சி நோய்களுக்கான இலக்கு சிகிச்சை மருந்துகளின் வளர்ச்சி சமீபத்திய ஆண்டுகளில் அதிக ஆர்வத்தை ஈர்த்துள்ளது. இந்த மதிப்பாய்வில், கீப்1 (கெல்ச் போன்ற ECH தொடர்புடைய புரதம்)/Nrf2 (NF-E2 p45-தொடர்புடைய காரணி 2)/ARE (ஆக்ஸிடன்ட் ரெஸ்பான்ஸ் உறுப்பு) அழற்சியின் சிக்னலிங் பாதை பற்றிய ஆராய்ச்சியை சுருக்கமாகக் கூறுகிறோம்.
Nrf2 இன் கட்டமைப்பு மற்றும் ஒழுங்குமுறை
Keap1-சார்ந்த Nrf2 ஒழுங்குமுறை
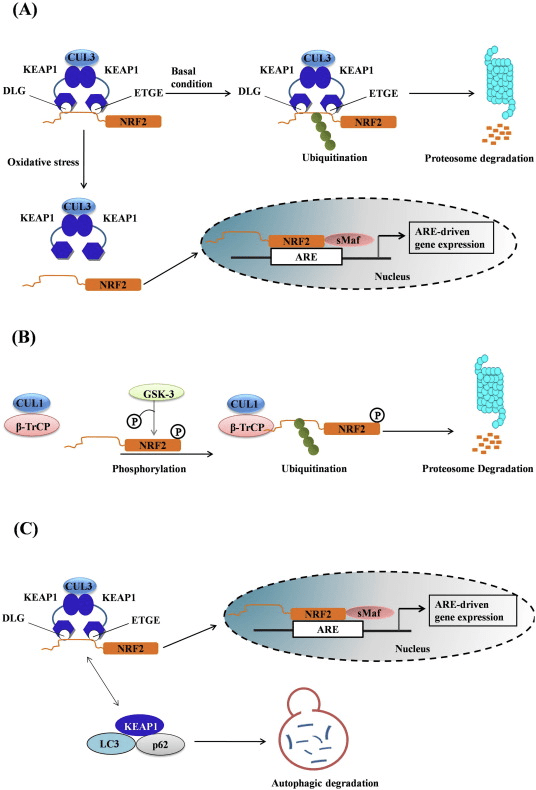
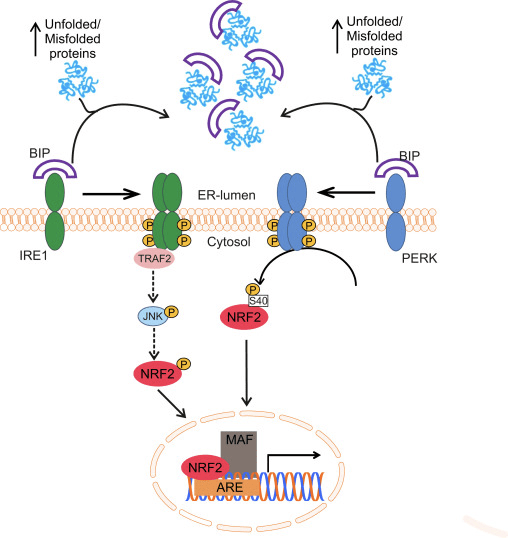
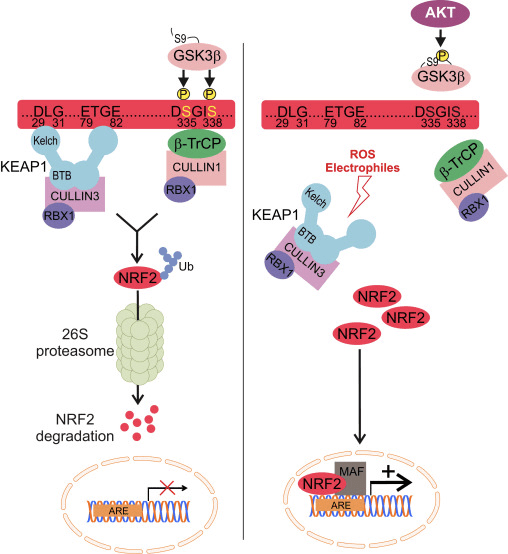
Nrf2 என்பது கேப் என் காலர் (CNC) துணைக் குடும்பத்தைச் சேர்ந்தது மற்றும் ஏழு செயல்பாட்டுக் களங்களை உள்ளடக்கியது, Neh (Nrf2-ECH ஹோமோலஜி) 1 முதல் Neh7 [9], [10]. Neh1 என்பது ஒரு CNC-bZIP டொமைன் ஆகும், இது Nrf2 ஐ சிறிய மஸ்குலோஅபோனியூரோடிக் ஃபைப்ரோசர்கோமா (மாஃப்) புரதம், டிஎன்ஏ மற்றும் பிற டிரான்ஸ்கிரிப்ஷன் பார்ட்னர்களுடன் ஹீட்டோரோடைமரைஸ் செய்ய அனுமதிக்கிறது, அத்துடன் ubiquitin-conjugating enzyme UbcM2 [11], [12]. Neh2 ஆனது DLG மற்றும் ETGE எனப்படும் இரண்டு முக்கியமான மையக்கருத்துக்களைக் கொண்டுள்ளது, அவை Nrf2 மற்றும் அதன் எதிர்மறை சீராக்கி Keap1 [13], [14] ஆகியவற்றுக்கு இடையேயான தொடர்புக்கு அவசியமானவை.
Keap1 என்பது குலின் அடிப்படையிலான E3 ubiquitin ligaseக்கான அடி மூலக்கூறு அடாப்டர் ஆகும், இது Nrf2 இன் டிரான்ஸ்கிரிப்ஷனல் செயல்பாட்டைத் தடுக்கிறது. Keap15 ஹோமோடைமரின் KELCH டொமைன்கள் சைட்டோசோலில் உள்ள Nrf16-Neh17 டொமைனின் DLG மற்றும் ETGE மையக்கருங்களுடன் பிணைக்கப்படுகின்றன, அங்கு ETGE அதிக தொடர்புடன் ஒரு கீலாக செயல்படுகிறது மற்றும் DLG ஒரு தாழ்ப்பாளாக செயல்படுகிறது [1]. ஆக்ஸிஜனேற்ற அழுத்தத்தின் கீழ் அல்லது Nrf2 ஆக்டிவேட்டர்களின் வெளிப்பாட்டின் போது, Keap2 சிஸ்டைன் எச்சங்களின் தியோல் மாற்றத்தின் காரணமாக Nrf18 ஆனது Keap2 பிணைப்பிலிருந்து பிரிகிறது, இது இறுதியில் Nrf2 எங்கும் பரவுதல் மற்றும் புரோட்டீசோமால் சிதைவைத் தடுக்கிறது [1]. பின்னர் Nrf1 கருவுக்குள் இடமாற்றம் செய்கிறது, சிறிய மாஃப் புரதங்களுடன் ஹீட்டோரோடைமரைஸ் செய்கிறது, மேலும் ARE பேட்டரி ஜீன்களை மாற்றுகிறது (படம் 2A). CHD19 (குரோமோ-ஏடிபேஸ்/ஹெலிகேஸ் டிஎன்ஏ பிணைப்பு புரதம்) [2] எனப்படும் டிரான்ஸ்கிரிப்ஷன் கோ-ஆக்டிவேட்டருடன் தொடர்புகொள்வதன் மூலம் Neh1 இன் கார்பாக்சி-டெர்மினல் டிரான்ஸ்கிரிப்ஷன் டொமைனாக செயல்படுகிறது. Neh3 மற்றும் Neh6 ஆகியவை பரிமாற்றக் களங்களாகவும் செயல்படுகின்றன, ஆனால் CBP (cAMP-response-element-binding protein-binding protein) [20] எனப்படும் மற்றொரு டிரான்ஸ்கிரிப்ஷனல் கோ-ஆக்டிவேட்டருடன் பிணைக்கப்படுகின்றன. மேலும், Neh4 மற்றும் Neh5 ஆகியவை அணுசக்தி இணைப்பான் RAC21/AIB4/SRC-5 உடன் தொடர்பு கொள்கின்றன, இது மேம்படுத்தப்பட்ட Nrf3-இலக்கு ARE மரபணு வெளிப்பாட்டிற்கு வழிவகுக்கிறது [1]. Neh3 ஆனது ரெடாக்ஸ்-சென்சிட்டிவ் அணு-ஏற்றுமதி சமிக்ஞையைக் கொண்டுள்ளது, இது Nrf2 [22] இன் ஒழுங்குமுறை மற்றும் செல்லுலார் உள்ளூர்மயமாக்கலுக்கு முக்கியமானது.
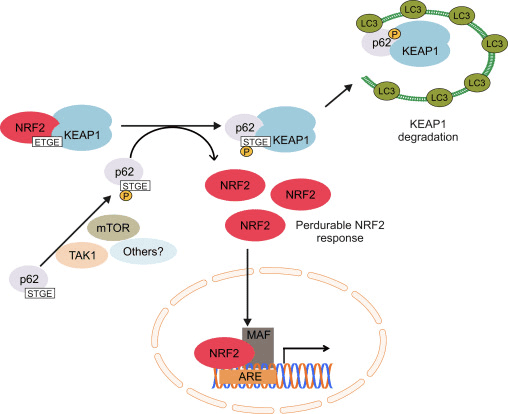
படம் 1 Keap1-சார்ந்த மற்றும் Nrf2 இன் சுயாதீன ஒழுங்குமுறை. (A) அடிப்படை நிலைமைகளின் கீழ், Nrf2 அதன் இரண்டு மையக்கருத்துகளால் (ETGE மற்றும் DLG) Keap1 உடன் பிரிக்கப்படுகிறது, இது CUL3-மத்தியஸ்தம் கொண்ட எங்கும் பரவுவதற்கு வழிவகுக்கிறது, அதைத் தொடர்ந்து புரோட்டீசோம் சிதைவு ஏற்படுகிறது. ஆக்ஸிஜனேற்ற அழுத்தத்தின் கீழ், Nrf2 Keap1 இலிருந்து பிரிந்து, கருவுக்கு இடமாற்றம் செய்து ARE-ஜீன் பேட்டரியை செயல்படுத்துகிறது. (B) GSK3 பாஸ்போரிலேட்டுகள் Nrf2 மற்றும் இது CUL2-மத்தியஸ்தம் செய்யப்பட்ட எங்கும் பரவுதல் மற்றும் அடுத்தடுத்த புரோட்டீசோம் சிதைவுக்கு ?-TrCP மூலம் Nrf1 ஐ அங்கீகரிக்க உதவுகிறது. (C) p62 ஆனது Keap1 உடன் வரிசைப்படுத்தப்பட்டு, அதன் தன்னியக்க சிதைவு, Nrf2 இன் விடுதலை மற்றும் அதிகரித்த Nrf2 சமிக்ஞைக்கு வழிவகுக்கிறது.
Keap1-சுயாதீன Nrf2 ஒழுங்குமுறை
வளர்ந்து வரும் சான்றுகள் Keap2 இலிருந்து சுயாதீனமான Nrf1 ஒழுங்குமுறையின் ஒரு புதிய வழிமுறையை வெளிப்படுத்தியுள்ளன. Nrf6 இன் செரீன் நிறைந்த Neh2 டொமைன் இந்த ஒழுங்குமுறையில் அதன் இரண்டு மையக்கருத்துக்களுடன் (DSGIS மற்றும் DSAPGS) ?-டிரான்ஸ்டூசின் ரிபீட்-கொண்ட புரதத்துடன் (?-TrCP) பிணைப்பதன் மூலம் முக்கிய பங்கு வகிக்கிறது [24]. ?-TrCP என்பது Skp1′Cul1′Rbx1/Roc1 ubiquitin ligase complexக்கான அடி மூலக்கூறு ஏற்பியாகும், இது Nrf2 ஐ எங்கும் பரவுதல் மற்றும் புரோட்டீசோமால் சிதைவுக்கு இலக்காகக் கொண்டுள்ளது. கிளைகோஜன் சின்தேஸ் கைனேஸ்-3 என்பது Keap1-சுயாதீனமான Nrf2 உறுதிப்படுத்தல் மற்றும் ஒழுங்குபடுத்தலில் ஈடுபட்டுள்ள ஒரு முக்கியமான புரதமாகும்; இது Nrf2 ஐ ?-TrCP மற்றும் அதைத் தொடர்ந்து புரதச் சிதைவு [6] (படம் 2B) மூலம் Nrf25 ஐ அங்கீகரிப்பதற்கு வசதியாக Neh1 டொமைனில் NrfXNUMX ஐ பாஸ்போரிலேட் செய்கிறது.
மற்ற Nrf2 கட்டுப்பாட்டாளர்கள்
மற்றொரு ஆதாரம் p62-சார்ந்த Nrf2 செயல்பாட்டின் நியமனமற்ற பாதையை வெளிப்படுத்தியுள்ளது, இதில் p62 Keap1 ஐ தன்னியக்க சிதைவுக்கு சீக்வெஸ்டர்கள் செய்கிறது, இது இறுதியில் Nrf2 இன் நிலைப்படுத்தலுக்கும் Nrf2-சார்ந்த மரபணுக்களின் பரிமாற்றத்திற்கும் வழிவகுக்கிறது [26], [27], [ 28], [29] (படம் 1C).
Nrf2 செயல்பாட்டைக் கட்டுப்படுத்துவதில் பல மைஆர்என்ஏக்கள் முக்கியப் பங்கு வகிக்கின்றன என்று ஆதாரங்களைக் குவிப்பது தெரிவிக்கிறது [30]. சங்கோகோயா மற்றும் பலர். [31] miR-144 நேரடியாக லிம்போபிளாஸ்ட் K2 செல் லைன், முதன்மை மனித எரித்ராய்டு ப்ரோஜெனிட்டர் செல்கள் மற்றும் அரிவாள்-செல் நோய் ரெட்டிகுலோசைட்டுகளில் Nrf562 செயல்பாட்டைக் குறைக்கிறது என்பதை நிரூபித்தது. மனித மார்பக எபிடெலியல் செல்களில் மற்றொரு சுவாரஸ்யமான ஆய்வு, கீப்28-சுயாதீன பொறிமுறையின் மூலம் miR-2 Nrf1 ஐத் தடுக்கிறது என்பதை நிரூபித்தது [32]. இதேபோல், miR-153, miR-27a, miR-142-5p, மற்றும் miR144 போன்ற miRNAகள் நியூரானல் SH-SY2Y செல் வரிசையில் Nrf5 வெளிப்பாட்டைக் குறைக்கின்றன [33]. சிங் மற்றும் பலர். [34] miR-93 இன் எக்டோபிக் வெளிப்பாடு Nrf2-ஒழுங்குபடுத்தப்பட்ட மரபணுக்களின் வெளிப்பாட்டை 17?-எஸ்ட்ராடியோல் (E2)-தூண்டப்பட்ட எலி மாதிரியான பாலூட்டி புற்றுநோயை உருவாக்குகிறது என்பதை நிரூபித்தது.
எங்கள் ஆய்வகத்தின் சமீபத்திய கண்டுபிடிப்பு, ரெட்டினோயிக் எக்ஸ் ரிசெப்டர் ஆல்பா (ஆர்எக்ஸ்ஆர்?) எனப்படும் என்ஆர்எஃப்2 இன் எண்டோஜெனஸ் இன்ஹிபிட்டரை அடையாளம் கண்டுள்ளது. RXR? ஒரு அணுக்கரு ஏற்பி, அதன் DNA-பிணைப்பு டொமைன் (DBD) வழியாக Nrf7 இன் Neh2 டொமைனுடன் (அமினோ-அமில எச்சங்கள் 209–316) தொடர்பு கொள்கிறது, மேலும் குறிப்பாக அணுக்கருவில் Nrf2 செயல்பாட்டைத் தடுக்கிறது. மேலும், பெராக்ஸிசோம் ப்ரோலிஃபெரேட்டர்-ஆக்டிவேட்டட் ரிசெப்டர்-?, ஈஆர்?, ஈஸ்ட்ரோஜன் தொடர்பான ரிசெப்டர்-?, மற்றும் குளுக்கோகார்டிகாய்டு ரிசெப்டர்கள் போன்ற பிற அணுக்கரு ஏற்பிகள் Nrf2 செயல்பாட்டின் எண்டோஜெனஸ் தடுப்பான்களாகவும் தெரிவிக்கப்பட்டுள்ளன [9], [10].
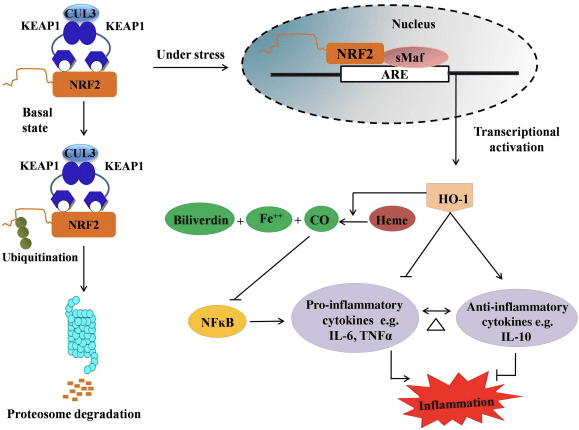
Nrf2/HO-1 அச்சின் அழற்சி எதிர்ப்பு பங்கு
HO-1 என்பது தூண்டக்கூடிய ஐசோஃபார்ம் மற்றும் விகிதத்தைக் கட்டுப்படுத்தும் என்சைம் ஆகும், இது ஹீம் கார்பன் மோனாக்சைடு (CO) மற்றும் இலவச இரும்பு மற்றும் பிலிவர்டின் பிலிரூபினாக சிதைவதை ஊக்குவிக்கிறது. புரோ-இன்ஃப்ளமேட்டரி ஃப்ரீ ஹீமின் நொதி சிதைவு மற்றும் CO மற்றும் பிலிரூபின் போன்ற அழற்சி எதிர்ப்பு சேர்மங்களின் உற்பத்தி HO-1 இன் பாதுகாப்பு விளைவுகளை பராமரிப்பதில் முக்கிய பங்கு வகிக்கிறது (படம். 2).
படம் 2 Nrf2/HO-1 பாதையின் மேலோட்டம். அடிப்படை நிலைமைகளின் கீழ், Nrf2 அதன் அடக்குமுறையான Keap1 உடன் பிணைக்கிறது, இது எங்கும் பரவுவதைத் தொடர்ந்து புரோட்டீசோம் சிதைவுக்கு வழிவகுக்கிறது. ஆக்ஸிஜனேற்ற அழுத்தத்தின் போது, இலவச Nrf2 அணுக்கருவிற்கு இடமாற்றம் செய்யப்படுகிறது, அங்கு அது சிறிய மாஃப் குடும்ப உறுப்பினர்களுடன் டைமரைஸ் செய்கிறது மற்றும் HO-1 போன்ற ARE மரபணுக்களுடன் பிணைக்கிறது. அதிக ஒழுங்குபடுத்தப்பட்ட HO-1 ஹீமை CO, பிலிரூபின் மற்றும் இலவச இரும்பாக மாற்றுகிறது. CO NF-?B பாதையின் தடுப்பானாக செயல்படுகிறது, இது அழற்சிக்கு சார்பான சைட்டோகைன்களின் வெளிப்பாடு குறைவதற்கு வழிவகுக்கிறது, அதே நேரத்தில் பிலிரூபின் ஆக்ஸிஜனேற்றியாகவும் செயல்படுகிறது. மேலும், HO-1 புரோஇன்ஃப்ளமேட்டரி சைட்டோகைன்களை நேரடியாகத் தடுக்கிறது மற்றும் அழற்சி எதிர்ப்பு சைட்டோகைன்களை செயல்படுத்துகிறது, இதனால் அழற்சி செயல்முறையை சமநிலைப்படுத்துகிறது.
Nrf2 mRNA மற்றும் புரத வெளிப்பாட்டை அதிகரிப்பதன் மூலம் HO-1 மரபணுவைத் தூண்டுகிறது மேலும் இது கிளாசிக் Nrf2 ஒழுங்குபடுத்தப்பட்ட மரபணுக்களில் ஒன்றாகும், இது பல விட்ரோ மற்றும் விவோ ஆய்வுகளில் பரவலாகப் பயன்படுத்தப்படுகிறது. HO-1 மற்றும் அதன் வளர்சிதை மாற்றங்கள் Nrf2 ஆல் மத்தியஸ்தம் செய்யப்பட்ட குறிப்பிடத்தக்க அழற்சி எதிர்ப்பு விளைவுகளைக் கொண்டுள்ளன என்பதை பல ஆய்வுகள் நிரூபித்துள்ளன. செயல்படுத்தப்பட்ட Nrf1 மூலம் மத்தியஸ்தம் செய்யப்படும் HO-2 வெளிப்பாட்டின் உயர்வானது NF?B சிக்னலைத் தடுப்பதற்கு வழிவகுக்கிறது, இது ஆண் ஸ்ப்ராக்-டாவ்லி எலி கல்லீரல் மாற்று அறுவை சிகிச்சை மாதிரியில் [35] குடல் மியூகோசல் காயம் மற்றும் இறுக்கமான-சந்தி செயலிழப்பை ஏற்படுத்துகிறது. Nrf2-சார்ந்த HO-1 வெளிப்பாட்டைக் கட்டுப்படுத்துவது H2O12 சைட்டோடாக்சிசிட்டி [2] இலிருந்து சுட்டி பெறப்பட்ட C2C36 மயோபிளாஸ்ட்களைப் பாதுகாக்கலாம். Nrf2-சார்ந்த HO-1 லிப்போபோலிசாக்கரைடு (LPS) - RAW264.7- அல்லது மவுஸ் பெரிட்டோனியல் மேக்ரோபேஜ்-பெறப்பட்ட நுரை செல் மேக்ரோபேஜ்களில் மத்தியஸ்த அழற்சி பதில்களில் தாக்கத்தை ஏற்படுத்துகிறது. Nrf2 செயல்பாடு நுரை செல் மேக்ரோபேஜ்களின் பினோடைப்பை நீக்குகிறது மற்றும் மேக்ரோபேஜ்களின் அளவற்ற வீக்கத்தைத் தடுக்கிறது, அவை பெருந்தமனி தடிப்புத் தோல் அழற்சியின் முன்னேற்றத்தில் முக்கிய பங்கு வகிக்கின்றன [37]. Nrf2/HO-1 அச்சு LPS தூண்டப்பட்ட மவுஸ் BV2 மைக்ரோகிளியல் செல்கள் மற்றும் மவுஸ் ஹிப்போகாம்பல் HT22 செல்களை பாதிக்கிறது, இது நியூரோ இன்ஃப்ளமேஷனில் தாக்கத்தை ஏற்படுத்துகிறது. மவுஸ் BV1 மைக்ரோகிளியல் செல்களில் Nrf2 பாதை வழியாக HO-2 வெளிப்பாட்டைக் கட்டுப்படுத்துதல், இது மவுஸ் ஹிப்போகாம்பல் HT22 செல்களின் உயிரணு இறப்பைப் பாதுகாக்கிறது [38]. மேலும், கோபால்ட்-அடிப்படையிலான கலப்பின மூலக்கூறுகள் (HYCOs) ஒரு Nrf2 தூண்டியை கார்பன் மோனாக்சைடு (CO) வெளியீட்டாளருடன் இணைக்கிறது, Nrf2/HO-1 வெளிப்பாட்டை அதிகரிக்கிறது, CO ஐ விடுவிக்கிறது மற்றும் விட்ரோவில் அழற்சி எதிர்ப்பு செயல்பாட்டைச் செய்கிறது. HYCO க்கள் திசு HO-1 ஐ அதிக-ஒழுங்குபடுத்துகின்றன மற்றும் விவோவில் நிர்வாகத்திற்குப் பிறகு இரத்தத்தில் CO ஐ வழங்குகின்றன, அழற்சி நிலைமைகளுக்கு எதிராக அவற்றின் சாத்தியமான பயன்பாட்டை ஆதரிக்கின்றன [39]. Nrf2/HO-1 முறைப்படுத்தல் டாரைன் குளோராமைன்களுடன் [40] சிகிச்சையளிக்கப்பட்ட முரைன் மேக்ரோபேஜ்களின் எஃபெரோசைடிக் செயல்பாட்டை அதிகரிப்பதன் மூலம் வீக்கத்தைக் குறைக்கிறது. ஒட்டுமொத்தமாக, மேலே விவரிக்கப்பட்ட சோதனை மாதிரிகள் அழற்சி எதிர்ப்பு செயல்பாட்டில் Nrf2/HO-1 அச்சு முக்கிய பங்கு வகிக்கிறது என்பதை வெளிப்படுத்தியது, இது அழற்சியுடன் தொடர்புடைய நோய்களில் Nrf2 ஒரு சிகிச்சை இலக்கு என்று பரிந்துரைக்கிறது.
கூடுதலாக, HO-1 இன் துணை தயாரிப்புகளான CO, பிலிரூபின், ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் செல் சேதத்தின் போது சக்திவாய்ந்த ஆக்ஸிஜனேற்றியாக செயல்படுகிறது [41], [42]; இது ஆட்டோ இம்யூன் என்செபலோமைலிடிஸ் மற்றும் ஹெபடைடிஸ் [43], [44] ஆகியவற்றை அடக்குகிறது; மேலும் இது iNOS மற்றும் NO [45], [46], [47] உருவாக்கத்தைத் தடுப்பதன் மூலம் எண்டோடாக்ஸிக் அதிர்ச்சியிலிருந்து எலிகள் மற்றும் எலிகளைப் பாதுகாக்கிறது. மேலும், பிலிரூபின் எண்டோடெலியல் செயல்பாடு மற்றும் செயலிழப்பைக் குறைக்கிறது [48]. சுவாரஸ்யமாக, பிலிரூபின் ஒட்டுதல் மூலக்கூறு-1 [49] வழியாக எண்டோடெலியல் லுகோசைட்டுகளின் பரிமாற்றத்தைக் குறைக்கிறது. இந்த குறிப்பிட்ட குறிப்புகள் HO-1 ஒரு சக்திவாய்ந்த அழற்சி எதிர்ப்பு முகவராக மட்டுமல்லாமல் அதன் வளர்சிதை மாற்றங்களையும் குறிக்கிறது.
அழற்சி மத்தியஸ்தர்கள் மற்றும் என்சைம்கள் Nrf2 ஆல் தடுக்கப்படுகின்றன
சைட்டோகைன்கள் மற்றும் கெமோக்கின்கள்
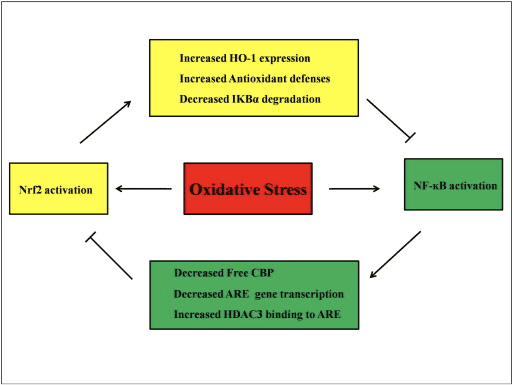
சைட்டோகைன்கள் குறைந்த மூலக்கூறு-எடை புரதங்கள் மற்றும் பல்வேறு உயிரணுக்களால் சுரக்கும் பாலிபெப்டைடுகள்; அவை உயிரணு வளர்ச்சி, வேறுபாடு மற்றும் நோயெதிர்ப்பு செயல்பாட்டை ஒழுங்குபடுத்துகின்றன, மேலும் வீக்கம் மற்றும் காயம்-குணப்படுத்துதலில் ஈடுபட்டுள்ளன. சைட்டோகைன்களில் இன்டர்லூகின்கள் (ILs), இன்டர்ஃபெரான்கள், கட்டி நெக்ரோசிஸ் காரணி (TNF), காலனி-தூண்டுதல் காரணி, கெமோக்கின்கள் மற்றும் வளர்ச்சி காரணிகள் ஆகியவை அடங்கும். சில சைட்டோகைன்கள் அழற்சி எதிர்ப்பு மத்தியஸ்தர்களாகக் கணக்கிடப்படுகின்றன, மற்றவை அழற்சி எதிர்ப்பு செயல்பாடுகளைக் கொண்டுள்ளன. ஆக்ஸிஜனேற்ற அழுத்தத்தின் வெளிப்பாடு சைட்டோகைன்களின் அதிகப்படியான உற்பத்தியில் விளைகிறது, இது இலக்கு செல்களில் ஆக்ஸிஜனேற்ற அழுத்தத்தை ஏற்படுத்துகிறது. ஆக்சிஜனேற்ற அழுத்தத்தால் NF-?B செயல்படுத்தப்படும்போது பல அழற்சிக்கு சார்பான சைட்டோகைன்கள் அதிகமாக உற்பத்தி செய்யப்படுகின்றன. மேலும், புரோ-இன்ஃப்ளமேட்டரி ஆக்ஸிஜனேற்ற அழுத்தமானது NF-?B மற்றும் சைட்டோகைன்களின் அதிக உற்பத்தியை மேலும் செயல்படுத்துகிறது. இந்த சுழற்சியை சீர்குலைப்பதில் Nrf2/ARE அமைப்பின் செயல்படுத்தல் முக்கிய பங்கு வகிக்கிறது. கெமோக்கின்கள் சிறிய சைட்டோகைன்களின் குடும்பமாகும், இதன் முக்கிய பங்கு அழற்சி உயிரணுக்களின் இடம்பெயர்வுக்கு வழிகாட்டுவதாகும். அவை முக்கியமாக லுகோசைட்டுகள், மோனோசைட்டுகள், நியூட்ரோபில்கள் மற்றும் பிற செயல்திறன் உயிரணுக்களுக்கான வேதியியல் கருவிகளாக செயல்படுகின்றன.
Nrf2 ஐ செயல்படுத்துவது IL-6 மற்றும் IL-1 உட்பட அழற்சிக்கு சார்பான சைட்டோகைன்களின் LPS-தூண்டப்பட்ட டிரான்ஸ்கிரிப்ஷனல் முறைப்படுத்தலைத் தடுக்கிறது என்று தெரிவிக்கப்பட்டுள்ளது. [50]. IL-1? மற்றும் IL-6 உற்பத்தி Nrf2?/? டெக்ஸ்ட்ரான் சல்பேட் தூண்டப்பட்ட பெருங்குடல் அழற்சி கொண்ட எலிகள் [51], [52]. Nrf2 கீழ்நிலை IL-17 மற்றும் பிற அழற்சி காரணிகளான Th1 மற்றும் Th17 ஆகியவற்றின் உற்பத்தியைத் தடுக்கிறது, மேலும் மல்டிபிள் ஸ்களீரோசிஸ், ஆட்டோ இம்யூன் என்செபாலிடிஸ் [53] இன் சோதனை மாதிரியில் நோய் செயல்முறையை அடக்குகிறது. Nrf2-சார்ந்த ஆன்டி-ஆக்ஸிடன்ட் மரபணுக்கள் HO-1, NQO-1, Gclc மற்றும் Gclm பிளாக் TNF-?, IL-6, மோனோசைட் கீமோ அட்ராக்டன்ட் புரதம்-1 (MCP1), மேக்ரோபேஜ் அழற்சி புரதம்-2 (MIP2) மற்றும் அழற்சி மத்தியஸ்தர்கள். ஆனால் Nrf2-நாக் அவுட் எலிகளின் விஷயத்தில், அழற்சி எதிர்ப்பு விளைவு ஏற்படாது [54]. LPS உடன் சிகிச்சையளிக்கப்பட்ட Nrf2-நாக் அவுட் எலிகளின் பெரிட்டோனியல் நியூட்ரோபில்கள், காட்டு-வகை (WT) செல்களைக் காட்டிலும் கணிசமாக அதிக அளவு சைட்டோகைன்கள் (TNF-? மற்றும் IL-6) மற்றும் கெமோக்கின்கள் (MCP1 மற்றும் MIP2) ஆகியவற்றைக் கொண்டுள்ளன [54]. விட்ரோவில், Nrf2 மரபணுவை மனித மற்றும் முயல் பெருநாடி மென்மையான தசை செல்களுக்கு மாற்றுவது MCP1 [8], [55] இன் சுரப்பை அடக்குகிறது, மேலும் Nrf2-சார்ந்த HO-1 வெளிப்பாடு TNF-?-தூண்டப்பட்ட NF-?B மற்றும் MCP-1 ஐ அடக்குகிறது. மனித தொப்புள் நரம்பு எண்டோடெலியல் செல்களில் சுரப்பு [56]. இந்த கண்டுபிடிப்புகள், அழற்சி தூண்டுதல்களுக்கு பதிலளிக்கும் விதமாக, Nrf2 சிக்னலை அதிகப்படுத்துவது அழற்சிக்கு சார்பான சைட்டோகைன்கள் மற்றும் கெமோக்கின்களின் அதிகப்படியான உற்பத்தியைத் தடுக்கிறது மற்றும் NF-?B இன் செயல்பாட்டைக் கட்டுப்படுத்துகிறது.
செல் ஒட்டுதல் மூலக்கூறுகள்
செல் ஒட்டுதல் மூலக்கூறுகள் (CAMs) செல்கள் அல்லது எக்ஸ்ட்ராசெல்லுலர் மேட்ரிக்ஸுடன் பிணைக்கும் புரதங்கள். செல் மேற்பரப்பில் அமைந்துள்ள, அவை செல் அங்கீகாரம், செல் செயல்படுத்தல், சமிக்ஞை கடத்துதல், பெருக்கம் மற்றும் வேறுபாடு ஆகியவற்றில் ஈடுபட்டுள்ளன. CAM களில், ICAM-1 மற்றும் VCAM-1 ஆகியவை இம்யூனோகுளோபுலின் சூப்பர் குடும்பத்தின் முக்கியமான உறுப்பினர்கள். ICAM-1 லுகோசைட் மற்றும் எண்டோடெலியல் செல் சவ்வுகளில் குறைந்த செறிவுகளில் உள்ளது. சைட்டோகைன் தூண்டுதலின் போது, செறிவு கணிசமாக அதிகரிக்கிறது. ICAM-1 ஐ IL-1 மற்றும் TNF ஆல் தூண்டலாம் மற்றும் வாஸ்குலர் எண்டோடெலியம், மேக்ரோபேஜ்கள் மற்றும் லிம்போசைட்டுகளால் வெளிப்படுத்தப்படுகிறது. இது லுகோசைட்டுகளில் காணப்படும் ஒரு ஏற்பியான இன்டெக்ரின் ஒரு தசைநார் ஆகும். ICAM-1-இன்டெக்ரின் பிரிட்ஜ் செயல்படுத்தப்படும்போது, லுகோசைட்டுகள் எண்டோடெலியல் செல்களுடன் பிணைந்து, பின்னர் சப்எண்டோதெலியல் திசுக்களுக்கு இடம்பெயர்கின்றன [57]. VCAM-1 லிம்போசைட்டுகள், மோனோசைட்டுகள், ஈசினோபில்கள் மற்றும் பாசோபில்களை வாஸ்குலர் எண்டோடெலியத்துடன் ஒட்டுவதற்கு மத்தியஸ்தம் செய்கிறது மற்றும் லுகோசைட் ஆட்சேர்ப்புக்கு பங்களிக்கிறது, இது இறுதியில் ஆக்ஸிஜனேற்ற அழுத்தத்தால் திசு சேதத்திற்கு வழிவகுக்கிறது. Nrf2 VCAM-1 [58] இன் ஊக்குவிப்பு செயல்பாட்டைத் தடுக்கிறது. Nrf2-ஒழுங்குபடுத்தப்பட்ட கீழ்நிலை மரபணு HO-1 ஆனது ஈ-செலக்டின் மற்றும் VCAM-1, எண்டோடெலியல் செல்களுடன் தொடர்புடைய ஒட்டுதல் மூலக்கூறுகளின் வெளிப்பாட்டை பாதிக்கலாம் [59]. CD-14, TREM1, SELE, SELP மற்றும் VCAM-1 போன்ற பல CAMகளின் நுரையீரல் வெளிப்பாடு Nrf2?/? Nrf2+/+ எலிகளை விட எலிகள் [60]. மனித பெருநாடி எண்டோடெலியல் செல்களில் உள்ள Nrf2 TNF-?-தூண்டப்பட்ட VCAM-1 வெளிப்பாட்டை அடக்குகிறது மற்றும் TNF-?-தூண்டப்பட்ட மோனோசைடிக் U937 செல் ஒட்டுதலில் தலையிடுகிறது [8]. Nrf2 இன் அதிகப்படியான வெளிப்பாடு TNF-?-தூண்டப்பட்ட VCAM-1 மரபணு வெளிப்பாட்டையும் மனித நுண்ணுயிர் எண்டோடெலியல் செல்களில் தடுக்கிறது [61]. இயற்கையாக நிகழும் ஆன்டிஆக்ஸிடன்ட் 3-ஹைட்ராக்ஸியாந்த்ரானிலிக் அமிலம் (HA), எல்-டிரிப்டோபான் வளர்சிதை மாற்றங்களில் ஒன்று, வீக்கம் அல்லது நோய்த்தொற்றின் போது கினுரேனைன் பாதை எனப்படும் வளர்சிதை மாற்ற பாதையில் விவோவில் உருவாகிறது, இது HO-1 வெளிப்பாட்டைத் தூண்டுகிறது மற்றும் மனித தொப்புளில் Nrf2 ஐத் தூண்டுகிறது. நரம்பு எண்டோடெலியல் செல்கள் (HUVECs). HA ஆல் தூண்டப்பட்ட Nrf2-சார்ந்த HO-1 வெளிப்பாடு MCP-1 சுரப்பு, VCAM-1 வெளிப்பாடு மற்றும் இரத்த நாளக் காயம் மற்றும் பெருந்தமனி தடிப்புத் தோல் அழற்சியுடன் தொடர்புடைய NF-kB செயல்படுத்தலைத் தடுக்கிறது [56]. பெருக்க எதிர்ப்பு மற்றும் அழற்சி எதிர்ப்பு செயற்கை சால்கோன் வழித்தோன்றல் 2?,4?,6?-ட்ரிஸ் (மெத்தாக்ஸிமெத்தாக்ஸி) சால்கோன் ICAM-1, அழற்சிக்கு சார்பான சைட்டோகைன் IL-1?, மற்றும் TNF-? டிரினிட்ரோபென்சீன் சல்போனிக் அமிலம் [62] மூலம் சிகிச்சையளிக்கப்பட்ட எலிகளிலிருந்து பெருங்குடல் திசுக்களில் வெளிப்பாடு. லைகோபீனுடன் சிகிச்சையளிக்கப்பட்ட மனித விழித்திரை நிறமி எபிடெலியல் செல்களில் TNF-?-தூண்டப்பட்ட ICAM-2 வெளிப்பாட்டைத் தடுக்கிறது [1]. இந்த ஆய்வுகள் அனைத்தும் அழற்சியின் செல்கள் வீக்கமடைந்த திசுக்களுக்கு இடம்பெயர்வு மற்றும் ஊடுருவலைக் கட்டுப்படுத்துவதன் மூலம் அழற்சி செயல்பாட்டில் Nrf63 முக்கிய பங்கு வகிக்கிறது என்று கூறுகின்றன.
மேட்ரிக்ஸ் மெட்டாலோபுரோட்டீனேஸ்கள் (MMPகள்)
எம்எம்பிகள் எக்ஸ்ட்ராசெல்லுலர் மேட்ரிக்ஸில் பரவலாக உள்ளன மற்றும் உயிரணு பெருக்கம், இடம்பெயர்வு, வேறுபாடு, காயம்-குணப்படுத்துதல், ஆஞ்சியோஜெனெசிஸ், அப்போப்டொசிஸ் மற்றும் கட்டி மெட்டாஸ்டாஸிஸ் போன்ற உடலியல் மற்றும் நோயியல் செயல்முறைகளில் ஈடுபட்டுள்ளன. Nrf2/HO-1 அச்சு மேக்ரோபேஜ்களில் MMP-9 ஐயும், மனித குடல் எபிடெலியல் செல்களில் MMP-7 ஐயும் தடுக்கிறது என்று தெரிவிக்கப்பட்டுள்ளது, மேலும் இது குடல் அழற்சி நோய் [62], [64] சிகிச்சையில் நன்மை பயக்கும். UV கதிர்வீச்சினால் தூண்டப்பட்ட தோல் சேதம் WT எலிகளை விட Nrf2-நாக் அவுட்டில் மிகவும் கடுமையானது மற்றும் MMP-9 நிலை கணிசமாக அதிகமாக உள்ளது, இது Nrf2 MMP-9 வெளிப்பாட்டைக் குறைக்கிறது என்பதைக் குறிக்கிறது. எனவே, Nrf2 புற ஊதா கதிர்வீச்சுக்கு எதிராக பாதுகாப்பாக கருதப்படுகிறது [65]. கட்டி செல் படையெடுப்பு மற்றும் வீக்கத்தில் MMP-9 இன் குறைக்கப்பட்ட டிரான்ஸ்கிரிப்ஷனல் செயல்படுத்தல் NF-kB சமிக்ஞை பாதையைத் தடுப்பதன் மூலம் கட்டுப்படுத்தப்படுகிறது என்றும் மற்றொரு ஆய்வு தெரிவிக்கிறது [66]. அதிர்ச்சிகரமான முதுகுத் தண்டு காயத்தில், MMP-9 [67] இன் mRNA அளவைக் கட்டுப்படுத்துவதில் NF-kB சமிக்ஞைப் பாதையும் பங்கேற்கிறது. எனவே, வீக்கத்தில் MMPகளின் கட்டுப்பாடு நேரடியாக Nrf2 பாதையால் அல்லது மறைமுகமாக Nrf2-பாதிக்கப்பட்ட NF-?B பாதை மூலம் பாதிக்கப்படுகிறது.
சைக்ளோஆக்சிஜனேஸ்-2 (COX2) மற்றும் தூண்டக்கூடிய நைட்ரிக் ஆக்சைடு சின்தேஸ் (INOS)
Nrf2-நாக் அவுட் எலிகள் மீதான தொடர்ச்சியான சோதனைகள், அழற்சியில் அதன் முக்கிய பங்கை நிரூபித்துள்ளன மற்றும் COX-2 மற்றும் iNOS போன்ற அழற்சிக்கு சார்பான மரபணுக்களை ஒழுங்குபடுத்துகின்றன. முதல் முறையாக, கோர் மற்றும் பலர். Nrf2 இன் பெருங்குடல் திசுக்களில் COX-2 மற்றும் iNOS போன்ற அழற்சிக்கு சார்பான சைட்டோகைன்களின் வெளிப்பாடு அதிகரித்துள்ளதாக தெரிவிக்கப்பட்டுள்ளது. WT Nrf2+/+ எலிகளுடன் ஒப்பிடும்போது, Nrf2 அவற்றின் செயல்பாட்டை அடக்குகிறது [51]. சிலுவை காய்கறிகளில் உள்ள நன்கு அறியப்பட்ட Nrf2 ஆக்டிவேட்டர்களில் ஒன்றான சல்ஃபோராபேன் உடனான முன் சிகிச்சை பற்றிய மற்றொரு அறிக்கை, mRNA இரண்டிலும் TNF-?, IL-1?, COX-2 மற்றும் iNOS ஆகியவற்றின் வெளிப்பாட்டைத் தடுப்பதன் மூலம் அதன் அழற்சி எதிர்ப்பு விளைவை நிரூபித்தது. மற்றும் Nrf2+/+ எலிகளிலிருந்து முதன்மை பெரிட்டோனியல் மேக்ரோபேஜ்களில் உள்ள புரத அளவுகள் Nrf2?/? எலிகள் [68]. இதேபோல், எல்பிஎஸ்-தூண்டப்பட்ட அழற்சியுடன் கூடிய Nrf2-நாக் அவுட் எலிகளின் ஹிப்போகாம்பஸ், iNOS, IL-6 மற்றும் TNF- போன்ற அழற்சி குறிப்பான்களின் அதிக வெளிப்பாட்டைக் காட்டுகிறது? WT எலிகளை விட [69]. அதேபோல், Nrf2-நாக் அவுட் எலிகள் 1-மெத்தில்-4-பீனைல்-1,2,3,6-டெட்ராஹைட்ரோபிரைடின் மூலம் தூண்டப்பட்ட ஆக்ஸிஜனேற்ற அழுத்தத்திற்கு அதிக உணர்திறன் கொண்டவை, மேலும் COX-2, iNOS போன்ற அழற்சி குறிப்பான்களின் அதிகரித்த mRNA மற்றும் புரத அளவுகளைக் காட்டுகின்றன. , IL-6, மற்றும் TNF-? [70]. மேலும், Nrf2 இலிருந்து கல்லீரல்?/? மெத்தியோனைன் மற்றும் கோலின்-குறைபாடுள்ள உணவில் சவால் செய்யப்பட்ட எலிகள் காக்ஸ்5 இன் ~ 2 மடங்கு அதிக mRNA வெளிப்பாடு மற்றும் iNOS ஐ ஒரே உணவில் உள்ள WT எலிகளை விட, Nrf2 [71] இன் அழற்சி எதிர்ப்புப் பங்கைக் குறிக்கிறது. சமீபத்தில், கிம் மற்றும் பலர். பிவி2 கலங்களில் Nrf2 சிக்னலிங் மூலம் iNOS இன் வெளிப்பாட்டைக் குறைப்பதன் மூலம் பைட்டோகெமிக்கல் எத்தில் பைருவேட் அதன் அழற்சி எதிர்ப்பு மற்றும் ஆக்ஸிஜனேற்ற விளைவுகளைச் செலுத்துகிறது என்பதை நிரூபித்தது. எத்தில் பைருவேட் Nrf2 இன் அணுக்கரு இடமாற்றத்தைத் தூண்டுகிறது, இது இறுதியில் p65 மற்றும் p300 இடையேயான தொடர்புகளைத் தடுக்கிறது, இது iNOS இன் வெளிப்பாடு குறைவதற்கு வழிவகுக்கிறது [72]. மேலும், கார்பசோல் அனலாக் LCY-2-CHO Nrf2 ஐ செயல்படுத்துகிறது மற்றும் அதன் அணுக்கரு இடமாற்றத்தை ஏற்படுத்துகிறது, இது எலி பெருநாடி வாஸ்குலர் மென்மையான தசை செல்களில் COX2 மற்றும் iNOS வெளிப்பாடு [73] அடக்குவதற்கு வழிவகுக்கிறது.
NLRP2 iIflammasome செயல்பாடுகளை ஒழுங்குபடுத்துவதில் Nrf3 இன் முரண்பாடான பங்கு
NLR குடும்பம், 3 (NLRP3) அழற்சியைக் கொண்ட பைரின் டொமைன் என்பது ஒரு மல்டிபுரோட்டீன் காம்ப்ளக்ஸ் ஆகும், இது நோய்க்கிருமி அங்கீகாரம் ஏற்பியாக (PRR) செயல்படுகிறது மற்றும் நோய்க்கிருமி-தொடர்புடைய மூலக்கூறு வடிவங்கள் (PAMPகள்), சேதம்- போன்ற நுண்ணுயிர், ஆக்ஸிஜனேற்ற அழுத்த சமிக்ஞைகளின் பரவலானதை அங்கீகரிக்கிறது. தொடர்புடைய மூலக்கூறு மாதிரி மூலக்கூறுகள் (DAMPகள்) மற்றும் ROS [74]. செயல்படுத்தப்பட்ட NLRP3 அழற்சியானது காஸ்பேஸ்-1 இன் பிளவு மற்றும் அழற்சிக்கு சார்பான சைட்டோகைன் இன்டர்லூகின்-1 இன் சுரப்பை மத்தியஸ்தம் செய்கிறது? (IL-1?) இது இறுதியில் பைரோப்டோசிஸ் எனப்படும் உயிரணு இறப்பின் செயல்முறையைத் தூண்டுகிறது, இது பரவலான நோய்க்கிருமிகளுக்கு எதிராக ஹோஸ்ட்களைப் பாதுகாக்கிறது [75]. இருப்பினும், அழற்சியின் பிறழ்ந்த செயல்பாடானது, டிரான்ஸ்மிசிபிள் ஸ்பாங்கிஃபார்ம் என்செபலோபதிஸ், அல்சைமர் நோய், பார்கின்சன் நோய் மற்றும் வகை 2 நீரிழிவு நோய் [76], புற்றுநோய் [77], கீல்வாதம் மற்றும் பெருந்தமனி தடிப்பு [78] போன்ற புரத தவறான மடிதல் நோய்களுடன் தொடர்புடையது.
ரோங் ஹு குழுவின் சமீபத்திய அவதானிப்பு, Nrf2 மற்றும் அழற்சியின் எதிர்மறையான ஒழுங்குமுறையுடன் Nrf2 ஆனது NLRP1 இன்ஃப்ளமேஸம் ஆக்டிவேஷன், காஸ்பேஸ்-3 பிளவு மற்றும் IL-1 ஆகியவற்றைத் தடுக்கும் NQO1 வெளிப்பாட்டைத் தூண்டுகிறது. மேக்ரோபேஜ்களில் தலைமுறை. மேலும், நன்கு அறியப்பட்ட Nrf2 ஆக்டிவேட்டர், tert-butylhydroquinone (tBHQ) ARE ஐ Nrf3-சார்ந்த முறையில் செயல்படுத்துவதன் மூலம் NLRP2 டிரான்ஸ்கிரிப்ஷனை எதிர்மறையாக ஒழுங்குபடுத்துகிறது [79]. மேலே உள்ள அவதானிப்புக்கு கூடுதலாக, அதே குழுவானது, Nrf2 அணுக்கரு இடமாற்றம் மற்றும் NLRP2 இன் இன்ஃப்ளமேஸம் அசெம்பிளி [3] தடுப்பதில் ஈடுபட்டுள்ள Nrf80 சிக்னலிங் பாதையை செயல்படுத்துவதன் மூலம் DSS-தூண்டப்பட்ட பெருங்குடல் அழற்சியைத் தடுக்கிறது.
இயற்கை மற்றும் செயற்கை சேர்மங்களைப் பயன்படுத்தி தொடர்ச்சியான சோதனைகள் NLRP2 இன் அழற்சி செயல்பாட்டில் Nrf3 இன் தடுப்பு விளைவை வெளிப்படுத்தியுள்ளன. உதாரணமாக, லூபஸ் நெஃப்ரிடிஸ் எலிகளில் epigallocatechin-3-gallate (EGCG) சிகிச்சையானது சிறுநீரக NLRP3 இன் இன்ஃப்ளமேஸம் ஆக்டிவேஷனைக் குறைப்பதைக் காட்டுகிறது, இது Nrf2 சிக்னலிங் பாதையால் மத்தியஸ்தம் செய்யப்படுகிறது [81]. அதேபோல், சிட்ரல் (3,7-டைமிதில்-2,6-ஆக்டாடியனல்), சீன மூலிகை மருந்தான லிட்சியா கியூபேபாவில் உள்ள ஒரு முக்கிய செயலில் உள்ள கலவை, துரிதப்படுத்தப்பட்ட மற்றும் தீவிரமான லூபஸ் நெஃப்ரிடிஸ் (ASLN) எலி மாதிரியில் NRf3 ஆக்ஸிஜனேற்ற சமிக்ஞை பாதை வழியாக NLRP2 அழற்சியை செயல்படுத்துவதைத் தடுக்கிறது. [82]. இதேபோல், Nrf2 பாதையை செயல்படுத்துவதன் மூலமும், ஆண் BALB/c எலிகளில் NLRP3 அழற்சி செயல்பாட்டைத் தடுப்பதன் மூலமும் LPS/GalN- தூண்டப்பட்ட கல்லீரல் காயத்திலிருந்து பயோசானின் பாதுகாக்கப்படுகிறது [83]. மேலும், மாங்கிஃபெரின் Nrf2 மற்றும் HO-1 இன் வெளிப்பாட்டை டோஸ்-சார்ந்த முறையில் கட்டுப்படுத்துவதாகக் காட்டப்பட்டது மற்றும் LPS/D-GalN- தூண்டப்பட்ட கல்லீரல் NLRP3, ASC, caspase-1, IL-1 ஆகியவற்றைத் தடுப்பதா? மற்றும் TNF-? வெளிப்பாடு [84].
Nrf3 ஆல் NLRP2 இன் எதிர்மறையான ஒழுங்குமுறை இருந்தபோதிலும், இது NLRP3 மற்றும் AIM2 அழற்சி செயல்பாட்டையும் செயல்படுத்துகிறது. ஹைடாவோ வென் மற்றும் சகாக்கள் அதை கண்டுபிடித்தனர், Nrf2 ?/? மவுஸ் மேக்ரோபேஜ்கள் NLRP3 மற்றும் AIM2 அழற்சியின் குறைபாடுள்ள செயல்பாட்டைக் காட்டுகின்றன, ஆனால் NLRC4 அழற்சியைக் காட்டவில்லை [85]. சுவாரஸ்யமாக, வீக்கத்துடன் தொடர்புடைய நோய்களின் பின்னணியில் Nrf2 இன் அறியப்படாத செயல்பாடுகளை இந்தக் கவனிப்பு சித்தரிக்கிறது; எனவே ஒரு சிகிச்சை இலக்காகக் கருதுவதற்கு முன், Nrf2 அழற்சி செயல்பாட்டைச் செயல்படுத்தும் பொறிமுறையை வெளிப்படுத்த மேலும் படிப்பது மிகவும் முக்கியம்.
Nrf2 மூலம் புரோ-இன்ஃப்ளமேட்டரி சைட்டோகைன் டிரான்ஸ்கிரிப்ஷனை அடக்குதல்
மவுஸ் மேக்ரோபேஜ்களில் குரோமாடின் இம்யூனோபிரெசிபிட்டேஷன் (சிஐபி)-சீக் மற்றும் சிஐபி-க்யூபிசிஆர் முடிவுகளை அடிப்படையாகக் கொண்ட மிக சமீபத்திய விசாரணையில், என்ஆர்எஃப்2 ஐஎல்-6 மற்றும் ஐஎல்-1 போன்ற அழற்சிக்கு எதிரான சைட்டோகைன்களின் ஊக்குவிப்பு பகுதிகளுடன் பிணைக்கிறது என்பதை வெளிப்படுத்தியது? மற்றும் RNA Pol II ஆட்சேர்ப்பைத் தடுக்கிறது. இதன் விளைவாக, RNA Pol II ஆல் IL-6 மற்றும் IL-1 இன் டிரான்ஸ்கிரிப்ஷனல் ஆக்டிவேஷனைச் செயல்படுத்த முடியவில்லை? இது இறுதியில் மரபணு வெளிப்பாட்டைத் தடுக்க வழிவகுக்கிறது. முதன்முறையாக, மசாயுகி யமமோட்டோவின் குழு புதிய வழிமுறையை வெளிப்படுத்தியது, இதன் மூலம் Nrf2 அதன் கீழ்நிலை மரபணுக்களை AREகள் மூலம் மாற்றுவது மட்டுமல்லாமல், RNA Pol II இன் ஆட்சேர்ப்பைத் தடுப்பதன் மூலம் ARE உடன் அல்லது இல்லாமலும் குறிப்பிட்ட மரபணுக்களின் டிரான்ஸ்கிரிப்ஷனல் செயல்பாட்டை அடக்குகிறது.
Nrf2 மற்றும் NF-?B பாதைகளுக்கு இடையே கிராஸ்டாக்
NF-?B என்பது கிட்டத்தட்ட அனைத்து வகையான விலங்கு உயிரணுக்களிலும் காணப்படும் DNA படியெடுத்தலுக்குப் பொறுப்பான ஒரு புரதச் சிக்கலானது மற்றும் வீக்கம், அப்போப்டொசிஸ், நோய் எதிர்ப்பு சக்தி, உயிரணு வளர்ச்சி மற்றும் வளர்ச்சி போன்ற பல்வேறு செயல்முறைகளில் ஈடுபட்டுள்ளது. p65, NF-?B குடும்பத்தின் Rel புரதம், ஒரு பரிமாற்ற டொமைனைக் கொண்டுள்ளது. ஆக்ஸிஜனேற்ற அழுத்தத்தின் போது, I?B கைனேஸ் (IKK) செயல்படுத்தப்பட்டு, I?B இன் பாஸ்போரிலேஷனை ஏற்படுத்துகிறது, இதன் விளைவாக NF-?B இன் வெளியீடு மற்றும் அணுக்கரு இடமாற்றம் ஏற்படுகிறது. NF-?B ஆனது IL-50, TNF-?, iNOS, IL-6 மற்றும் உள்செல்லுலார் ஒட்டுதல் COX-1 போன்ற அழற்சி-சார்பு மத்தியஸ்தர்களின் படியெடுத்தலை ஏற்படுத்துகிறது.
NF-?B இன் அசாதாரண ஒழுங்குமுறை முடக்கு வாதம், ஆஸ்துமா, அழற்சி குடல் நோய் மற்றும் ஹெலிகோபாக்டர் பைலோரி தொற்று-தூண்டப்பட்ட இரைப்பை அழற்சி [86] ஆகியவற்றுடன் இணைக்கப்பட்டுள்ளது. NF-kB செயல்பாடு Keapl/Nrf2/ARE சிக்னலிங் பாதையை முக்கியமாக மூன்று அம்சங்களில் பாதிக்கிறது என்று தற்போது கருதப்படுகிறது: முதலில், Keap1 IKK ஐக் குறைக்குமா? எங்கும் பரவுதல் மூலம், இதனால் NF-?B [87] செயல்பாட்டைத் தடுக்கிறது. இரண்டாவதாக, அழற்சி செயல்முறையானது சைக்ளோபென்டெனோன் புரோஸ்டாக்லாண்டின் 2d-PGJ15 இலிருந்து பெறப்பட்ட COX2 போன்ற அழற்சி மத்தியஸ்தர்களைத் தூண்டுகிறது, இது Keap1 உடன் வினைபுரியும் மற்றும் Nrf2 ஐ செயல்படுத்தும் ஒரு வலிமையான எலக்ட்ரோஃபைல் ஆகும், இதனால் ஒரே நேரத்தில் NF-kB செயல்பாட்டைத் தடுப்பதன் மூலம் மரபணு டிரான்ஸ்கிரிப்ஷனைத் தொடங்குகிறது [58], படம் 88 A, B). மூன்றாவதாக, NF-?B ஆனது போட்டி Nrf3 டிரான்ஸ்கிரிப்ஷனல் கோ-ஆக்டிவேட்டர் CBP [2], [89] (படம் 90 C, D) உடன் இணைக்க முடியும்.
படம் 3 Nrf2 மற்றும் NF-?B பாதைகளுக்கு இடையே உள்ள கிராஸ்டாக். (A) Keap1 ஆனது IKK ஐ CUL3-மத்தியஸ்தம் கொண்ட எங்கும் பரவுதல் மற்றும் புரோட்டீசோம் சிதைவுக்கு வழிநடத்துகிறது, இது இறுதியில் NF-?B பாஸ்போரிலேஷனைத் தடுக்க வழிவகுக்கிறது, மேலும் இந்த பொறிமுறையானது Keap2 உடன் Nrf1 மற்றும் IKK இன் போட்டிப் பிணைப்பாகவும் செயல்படுகிறது. (B) ஆக்ஸிஜனேற்ற அழுத்தம் IKK ஐ செயல்படுத்துகிறது, இது NF-?B ஐ பாஸ்போரிலேட் செய்கிறது, இது கருவுக்குள் அதன் இடமாற்றம் மற்றும் COX-2 போன்ற புரோஇன்ஃப்ளமேட்டரி சைட்டோகைன்களை செயல்படுத்துகிறது. 2d-PGJ15 என அழைக்கப்படும் COX-2 இன் முனைய தயாரிப்பு Nrf2 இன் தூண்டியாக செயல்படுகிறது, இது இறுதியில் ஆக்ஸிஜனேற்ற அழுத்தத்தை அடக்குவதற்கு வழிவகுக்கிறது. (C) Nrf2 அதன் டிரான்ஸ்கிரிப்ஷனல் கோஃபாக்டர் CBP உடன் சிறிய Maf மற்றும் பிற டிரான்ஸ்கிரிப்ஷனல் இயந்திரங்களுடன் இணைந்து ARE- இயக்கப்படும் மரபணு வெளிப்பாட்டைத் தொடங்குகிறது. (D) NF-?B CBP உடன் போட்டித்தன்மையுடன் பிணைக்கும்போது, Nrf2 உடன் CBP பிணைப்பைத் தடுக்கிறது, இது Nrf2 பரிமாற்றத்தைத் தடுக்கிறது.
கீழ்நிலை இலக்கு புரதங்களின் படியெடுத்தல் அல்லது செயல்பாட்டைக் கட்டுப்படுத்த Nrf2 மற்றும் NF-?B சமிக்ஞை பாதைகள் தொடர்பு கொள்கின்றன என்று கருதப்படுகிறது. இந்த அனுமானத்தை நியாயப்படுத்துவதில், Nrf2 மற்றும் NF-?B பாதைகளின் உறுப்பினர்களுக்கு இடையே நேரடி அல்லது மறைமுக செயல்படுத்தல் மற்றும் தடுப்பு ஏற்படுவதை பல எடுத்துக்காட்டுகள் காட்டுகின்றன (படம். 4). LPSக்கு பதிலளிக்கும் விதமாக, Nrf2 நாக் டவுன் NF-?B டிரான்ஸ்கிரிப்ஷனல் செயல்பாடு மற்றும் NF-?B-சார்ந்த மரபணு டிரான்ஸ்கிரிப்ஷனை கணிசமாக அதிகரிக்கிறது, Nrf2 NF-?B செயல்பாட்டைத் தடுக்கிறது [60], [91]. கூடுதலாக, Nrf2-சார்ந்த கீழ்நிலை HO-1 இன் அதிகரித்த வெளிப்பாடு NF-?B செயல்பாட்டைத் தடுக்கிறது. புரோஸ்டேட் புற்றுநோய் செல்கள் சுருக்கமாக ?-டோகோபெரில் சுசினேட்டிற்கு வெளிப்படும் போது, வைட்டமின் E இன் வழித்தோன்றல், HO-1 வெளிப்பாடு அதிகப்படுத்தப்படுகிறது. HO-1 இன் இறுதிப் பொருட்கள் NF-?B [92] இன் அணுக்கரு இடமாற்றத்தைத் தடுக்கின்றன. இவை விவோ ஆய்வுகளில் NRf2 எதிர்மறையாக NF-kB சமிக்ஞை பாதையை ஒழுங்குபடுத்துகிறது என்று கூறுகின்றன. LPS NF-?B DNA பிணைப்பு செயல்பாட்டைத் தூண்டுகிறது மற்றும் NF-?B இன் p65 துணைக்குழுவின் அளவு Nrf2 இன் நுரையீரலில் இருந்து அணுக்கரு சாற்றில் கணிசமாக அதிகமாக உள்ளது?/? WT எலிகளை விட, NF-?B செயல்படுத்தலில் Nrf2 இன் எதிர்மறையான பாத்திரத்தை பரிந்துரைக்கிறது. மேலும், Nrf2?/? மவுஸ் எம்ப்ரியோ ஃபைப்ரோபிளாஸ்ட்கள் எல்பிஎஸ் மற்றும் டிஎன்எஃப்-யுடன் சிகிச்சையளிக்கப்பட்டதா? IKK ஆக்டிவேஷனால் ஏற்படும் NF-?B செயல்படுத்தல் மற்றும் I?B-? சீரழிவு [60]. NRf2?/ இல் NF-?B டிஎன்ஏ-பிணைப்பு செயல்பாடு அதிகரிக்கும் போது சுவாச ஒத்திசைவு வைரஸ் அனுமதி கணிசமாகக் குறைக்கப்படுகிறது. WT எலிகளுடன் ஒப்பிடும்போது எலிகள் [93]. Nrf2?/ இல் பிரிஸ்டேன் தூண்டப்பட்ட லூபஸ் நெஃப்ரிடிஸ்? சல்ஃபோராஃபேனுடன் இணைந்து சிகிச்சையளிக்கப்பட்ட எலிகள் கடுமையான சிறுநீரக பாதிப்பு மற்றும் நோயியல் மாற்றங்கள் மற்றும் WT உடன் ஒப்பிடும்போது உயர்ந்த iNOS வெளிப்பாடு மற்றும் NF-?B செயல்படுத்தல் ஆகியவற்றைக் கொண்டுள்ளன, NRf2 NF-?B சமிக்ஞை பாதையைத் தடுப்பதன் மூலம் லூபஸ் நெஃப்ரிடிஸை மேம்படுத்துகிறது மற்றும் ROS [94] என்று பரிந்துரைக்கிறது. ]. NF-?B செயல்பாடு LPS மற்றும் TNF-? உடன் Nrf2 தூண்டியுடன் சிகிச்சை அளிக்கப்படும்போதும் ஏற்படுகிறது. எடுத்துக்காட்டாக, ஒரு செயற்கை சால்கோன் வழித்தோன்றல் TNF-?-தூண்டப்பட்ட NF-?B செயல்படுத்தலை நேரடியாகவும் மறைமுகமாகவும் மற்றும் ஓரளவு மனித குடல் எபிடெலியல் HT-1 செல்களில் HO-29 வெளிப்பாட்டின் தூண்டுதலின் மூலம் தடுக்கிறது [62]. F344 எலிகளை 3H-1,2-dithiole-3-thione (D3T) [95] கொண்டு சிகிச்சையளிக்கும் போது NF-?B இடமாற்றம் மற்றும் DNA-பிணைப்பு செயல்பாடு மற்றும் ஹெபடோசைட்டுகளில் iNOS வெளிப்பாட்டின் அடக்கம் ஆகியவை கண்டறியப்படுகின்றன. சல்ஃபோராபேன் மற்றும் எல்பிஎஸ் உடனான இணை சிகிச்சைக்குப் பிறகு, iNOS, COX-2 மற்றும் TNF- ஆகியவற்றின் LPS-தூண்டப்பட்ட வெளிப்பாடு? ரா 264.7 மேக்ரோபேஜ்கள் குறைக்கப்படுகின்றன, NF-?B டிஎன்ஏ பிணைப்பை [96] தடுப்பதன் மூலம் சல்ஃபோராபேன் அழற்சி எதிர்ப்பு செயல்பாட்டைக் கொண்டுள்ளது என்று பரிந்துரைக்கப்படுகிறது. Nrf2 மற்றும் NF-?B பாதைகளுக்கு இடையே உள்ள தொடர்பை விளக்க பல சோதனை ஆய்வுகள் செய்யப்பட்டாலும், முரண்பட்ட முடிவுகள் உள்ளன. Nrf2 மற்றும் NF-kB [97] இடையே நேர்மறை மற்றும் எதிர்மறை விதிமுறைகள் இரண்டும் பதிவாகியுள்ளன. பொதுவாக, வேதியியல் தடுப்பு எலக்ட்ரோஃபைல்கள் 3H-1,2-dithiole-3-thione, sulforapane மற்றும் Triterpenoid CDDO-Me ஆகியவை NF-kB மற்றும் அதன் குறைக்கப்பட்ட மரபணுக்களைத் தடுப்பதன் மூலம் Nrf2 ஐ செயல்படுத்துகின்றன [98], [99], [100]. இதற்கு நேர்மாறாக, ROS, LPS, ஓட்டம் வெட்டு அழுத்தம், ஆக்ஸிஜனேற்றப்பட்ட LDL மற்றும் சிகரெட் புகை போன்ற பல முகவர்கள் அல்லது நிலைமைகள் Nrf2 மற்றும் NF-kB செயல்பாட்டை அதிகரிப்பதாகக் காட்டப்பட்டுள்ளது [97]. கூடுதலாக, விவோ ஆய்வுகளில் NRf2 இலிருந்து தனிமைப்படுத்தப்பட்ட கல்லீரல்களில் NF-kB செயல்பாடு குறைந்துள்ளது என்று தெரியவந்துள்ளது. எலிகள் மற்றும் NF-?B பிணைப்பு செயல்பாடு Nrf2 இல் குறைவாக உள்ளதா?/? Nrf2+/+ எலிகளை விட [101]. இருப்பினும், அடினோவைரல் வெக்டார் Nrf2 உடன் சிகிச்சையளிக்கப்பட்ட மனித பெருநாடி எண்டோடெலியல் செல்கள் NF-?B இன் செயல்பாட்டை பாதிக்காமல் NF-?B கீழ்நிலை மரபணுக்களை தடுக்கிறது [8].
படம் 4 Nrf2 மற்றும் NF-?B இன் ஒழுங்குமுறை வளையம். Nrf2 பாதையானது I?B-யின் சிதைவைத் தடுப்பதன் மூலம் NF-?B செயல்படுத்தலைத் தடுக்கிறது. மேலும் HO-1 வெளிப்பாடு மற்றும் ஆக்ஸிஜனேற்ற பாதுகாப்புகளை அதிகரிக்கிறது, இது ROS மற்றும் நச்சுத்தன்மையை நீக்கும் இரசாயனங்களை நடுநிலையாக்குகிறது. இதன் விளைவாக, ROS-தொடர்புடைய NF-?B செயல்படுத்தல் ஒடுக்கப்பட்டது. அதேபோல், NF-?B-மத்தியஸ்த டிரான்ஸ்கிரிப்ஷன் Nrf2 செயல்பாட்டைக் குறைப்பதன் மூலம் குறைக்கிறதுஉள்ளீர்கள்CBP க்காக Nrf2 உடன் போட்டியிடுவதன் மூலம் மரபணு படியெடுத்தல் மற்றும் இலவச CREB பிணைப்பு புரதம். மேலும், NF-?B ஹிஸ்டோன் டீசெடைலேஸின் (HDAC3) ஆட்சேர்ப்பை ARE பிராந்தியத்திற்கு அதிகரிக்கிறது, எனவே Nrf2 டிரான்ஸ்கிரிப்ஷனல் செயல்படுத்தல் தடுக்கப்படுகிறது.
மனித உடலில் உள்ள உயிரணுக்களின் ஆக்ஸிஜனேற்ற திறனை மேம்படுத்துவதன் மூலம் எதிர்வினை ஆக்ஸிஜனேற்றத்தின் நச்சுத்தன்மையில் ஈடுபட்டுள்ள நொதிகள் மற்றும் மரபணுக்களின் வெளிப்பாட்டில் Nrf2 சிக்னலிங் பாதையின் செயல்படுத்தல் முக்கிய பங்கு வகிக்கிறது. இன்று பல ஆராய்ச்சி ஆய்வுகள் கிடைத்தாலும், Nrf2 செயல்படுத்தலில் உள்ள ஒழுங்குமுறை வழிமுறைகள் முழுமையாக புரிந்து கொள்ளப்படவில்லை. அழற்சியின் சிகிச்சையில் Nrf2 சமிக்ஞை பாதையின் சாத்தியமான பங்கும் கண்டறியப்பட்டுள்ளது. டாக்டர். அலெக்ஸ் ஜிமினெஸ் DC, CCST இன்சைட்
அழற்சி நோய்களில் Nrf2 இன் பங்கு
பல்வேறு அமைப்புகளை பாதிக்கும் அழற்சி நோய்களில் Nrf2 முக்கிய பங்கு வகிக்கிறது என்று vivo ஆய்வுகள் காட்டுகின்றன; இரைப்பை அழற்சி, பெருங்குடல் அழற்சி, கீல்வாதம், நிமோனியா, கல்லீரல் பாதிப்பு, இருதய நோய், நரம்பியக்கடத்தல் நோய் மற்றும் மூளை பாதிப்பு ஆகியவை இதில் அடங்கும். இந்த ஆய்வுகளில், Nrf2?/? WT விலங்குகளை விட விலங்குகள் வீக்கம் மற்றும் திசு சேதத்தின் கடுமையான அறிகுறிகளைக் காட்டின. எனவே, அழற்சி நோய்களில் Nrf2 சமிக்ஞை பாதை ஒரு பாதுகாப்பு விளைவைக் கொண்டுள்ளது என்று நம்பப்படுகிறது. போர்சின் கணைய எலாஸ்டேஸின் உள் மூச்சுக்குழாய் நிறுவல் நாள்பட்ட தடுப்பு நுரையீரல் நோயைத் தூண்டுகிறது, குறிப்பாக எம்பிஸிமா. Nrf2-குறைபாடுள்ள எலிகள் எம்பிஸிமாவுக்கு மிகவும் எளிதில் பாதிக்கப்படுகின்றன. நுரையீரல் காயத்திற்கு எதிரான மேக்ரோபேஜ் மத்தியஸ்த பாதுகாப்பு அமைப்பில் Nrf2 ஒரு முக்கிய கட்டுப்பாட்டாளராகக் கருதப்படுகிறது [102]. Nrf2-குறைபாடுள்ள எலிகள் புகையிலை புகை வெளிப்பாட்டால் 6 மாதங்களுக்கு தூண்டப்பட்ட மூச்சுக்குழாய் அழற்சியின் அதிகரிப்பு, அல்வியோலியில் ஆக்ஸிஜனேற்ற அழுத்த குறிப்பான்களின் ஒழுங்குபடுத்தப்பட்ட வெளிப்பாடு மற்றும் அதிகரித்த அல்வியோலர் செப்டல் செல் அப்போப்டொசிஸ் ஆகியவற்றைக் காட்டுகின்றன. மரபணுக்கள் [2], [102]. Nrf2 இடையூறுடன், ஒவ்வாமை-மத்தியஸ்த காற்றுப்பாதை அழற்சி மற்றும் ஓவல்புமின் வளாகத்தைப் பயன்படுத்தும் ஆஸ்துமா அதிகரித்த காற்றுப்பாதை அழற்சி, காற்றுப்பாதை அதி-வினைத்திறன், கோபட் செல்களின் ஹைப்பர் பிளாசியா மற்றும் மூச்சுக்குழாய் அழற்சி மற்றும் ஸ்ப்ளெனோசைட்டுகளில் அதிக அளவு Th2 ஐக் காட்டுகிறது. , சளி மிகை சுரப்பு, மற்றும் காற்றுப்பாதை அதி-வினைத்திறன் மற்றும் ஆஸ்துமாவின் வளர்ச்சியைத் தடுக்கும் பல ஆக்ஸிஜனேற்ற மரபணுக்களைத் தூண்டுகிறது [2]. ப்ளூரல் குழிக்குள் கராஜீனன் உட்செலுத்துதல் ப்ளூரிசியைத் தூண்டுகிறது, மேலும் Nrf15 அழற்சி உயிரணுக்களில் 2d-PGJ2 திரட்சியானது மவுஸ் பெரிட்டோனியல் மேக்ரோபேஜ்களுடன் மட்டுப்படுத்தப்பட்டுள்ளது. அழற்சியின் ஆரம்ப கட்டத்தில், 15d-PGJ2 Nrf2 ஐ செயல்படுத்துகிறது மற்றும் HO-1 மற்றும் PrxI இன் தூண்டல் மூலம் அழற்சி செயல்முறையை ஒழுங்குபடுத்துகிறது. 2d-PGJ15 [2] உற்பத்தியின் மூலம் ஆரம்ப கட்டத்தில் COX-105 அழற்சி எதிர்ப்பு விளைவைக் கொண்டுள்ளது என்றும் ஒரு ஆய்வு பரிந்துரைத்தது. 1 வாரத்திற்கு 1% டெக்ஸ்ட்ரான் சல்பேட் சோடியத்தின் வாய்வழி நிர்வாகம் பெருங்குடல் அழற்சியைத் தூண்டுகிறது, இதில் ஹிஸ்டோலாஜிக்கல் மாற்றங்களுடன் தொடர்புடைய கிரிப்ட்களின் சுருக்கம் மற்றும் பெருங்குடல் திசுக்களில் அழற்சி செல்கள் ஊடுருவல் ஆகியவை அடங்கும். பெருங்குடல் அழற்சியில் குடல் ஒருமைப்பாட்டைப் பாதுகாக்க, அழற்சிக்கு சார்பான சைட்டோகைன்களை ஒழுங்குபடுத்துவதன் மூலமும், இரண்டாம் கட்ட நச்சு நீக்கும் நொதிகளைத் தூண்டுவதன் மூலமும் Nrf2 முக்கியப் பங்காற்ற முடியும் [51]. LPS-தூண்டப்பட்ட நுரையீரல் செப்சிஸின் Nrf2-நாக் அவுட் மவுஸ் மாதிரியில், NF-?B செயல்பாடு COX-2, IL-113, IL-6 மற்றும் TNF போன்ற அழற்சி சைட்டோகைன்களின் செல்வாக்கைக் கட்டுப்படுத்துகிறது? வீக்கத்தைத் தொடங்குவதற்கும் ஊக்குவிப்பதற்கும் அவசியமானவை [60]. இந்த அழற்சி காரணிகளை ஒழுங்குபடுத்துவதன் மூலம் Nrf2 அழற்சி சேதத்தை குறைக்கிறது. கடுமையான அழற்சியின் இந்த மாதிரிகளில், Nrf2 சிக்னலிங் பாதையால் ஆக்ஸிஜனேற்ற நொதிகள், அழற்சிக்கு சார்பான சைட்டோகைன்கள் மற்றும் மத்தியஸ்தர்களின் அதிகரித்த கட்டுப்பாடு WT விலங்குகளில் ஏற்படும் அழற்சி காயத்தைக் குறைக்கிறது. சுவாரஸ்யமாக, இது Nrf2-நாக் அவுட் எலிகளிலும் பதிவாகியுள்ளது, இதில் WT எலிகளுடன் ஒப்பிடும்போது அறிகுறிகள் குறிப்பிடத்தக்க அளவில் அதிகரிக்கின்றன.
Nrf2-சார்ந்த அழற்சி எதிர்ப்பு மருந்துகள் பற்றிய ஆராய்ச்சி
சுருக்கமாக, அழற்சியின் பல பகுதிகளில் Nrf2 சமிக்ஞை பாதை ஒரு ஒழுங்குமுறைப் பாத்திரத்தை வகிக்கிறது என்பதைக் காட்டும் சோதனைகளைப் பற்றி நாங்கள் விவாதித்தோம், எனவே அழற்சி நோய்களுக்கான சிகிச்சைக்கு Nrf2-சார்ந்த அழற்சி எதிர்ப்பு முகவர்கள் முக்கியமானவை.
தாவரங்கள் Nrf2 டிரான்ஸ்கிரிப்ஷன் காரணியை செயல்படுத்தும் சேர்மங்களின் அசாதாரண வளமான ஆதாரங்களாக உள்ளன, இது சைட்டோபுரோடெக்டிவ் மரபணுக்களின் மேல்-ஒழுங்குமுறைக்கு வழிவகுக்கிறது. சமீபத்தில், பல்வேறு அழற்சி எதிர்ப்பு முகவர்களின் விளைவுகளை ஆராய பல ஆய்வுகள் நடத்தப்பட்டன, பெரும்பாலும் தாவர தோற்றம். எடுத்துக்காட்டாக, குர்குமின் என்பது மஞ்சளின் செயலில் உள்ள மூலப்பொருள் மற்றும் இஞ்சியில் சிறிய அளவில் காணப்படுகிறது; ஐசோதியோசயனேட்டுகள், குறிப்பாக ஃபெனிலிசோதியோசயனேட்டுகள் ப்ரோக்கோலி, செலரி மற்றும் பிற காய்கறிகளிலிருந்து பெறப்படுகின்றன; மற்றும் அந்தோசயினின்கள் பெர்ரி மற்றும் திராட்சை [124]. இந்த அனைத்து முகவர்களும் நல்ல ஆக்ஸிஜனேற்றிகள் மட்டுமல்ல, Nrf2 தூண்டல் [125], [126] மூலம் சக்திவாய்ந்த அழற்சி எதிர்ப்பு விளைவுகளையும் கொண்டிருப்பதாக ஆய்வுகள் காட்டுகின்றன. எனவே, தாவர சாற்றில் இருந்து புதிய அழற்சி எதிர்ப்பு Nrf2 ஆக்டிவேட்டர்களின் உருவாக்கம் மருத்துவ ஆராய்ச்சியில் அதிக ஆர்வத்தை ஈர்த்துள்ளது.
சமீபத்திய ஆண்டுகளில், இந்த சேர்மங்களின் செயல்களை உறுதிப்படுத்த பல விலங்கு பரிசோதனைகள் நடத்தப்பட்டுள்ளன. ஆர்ட்சுனேட் முக்கியமாக கடுமையான மலேரியா, பெருமூளை மலேரியா மற்றும் ருமேடிக் ஆட்டோ இம்யூன் நோய்களுக்கு பயன்படுத்தப்படுகிறது; செப்டிக் நுரையீரல் காயத்திலும் இது பயனுள்ளதாக இருக்கும். ஆர்ட்டசுனேட் Nrf2 மற்றும் HO-1 வெளிப்பாட்டை செயல்படுத்துகிறது, மேலும் பிந்தையது வீக்கத்தைத் தடுக்க அழற்சிக்கு சார்பான சைட்டோகைன்கள் மற்றும் லுகோசைட்டுகளின் ஊடுருவலைக் குறைக்கிறது [127]. ஓரிசா சாடிவா அரிசியின் தோலிலிருந்து பிரித்தெடுக்கப்படும் ஐசோவிடெக்சின், அழற்சி எதிர்ப்பு மற்றும் ஆக்ஸிஜனேற்ற பண்புகளைக் கொண்டிருப்பதாகக் கருதப்படுகிறது; Nrf2/HO-1 பாதையை செயல்படுத்தி MAPK மற்றும் NF-?B [128] ஆகியவற்றை செயல்படுத்துவதன் மூலம் LPS-தூண்டப்பட்ட கடுமையான நுரையீரல் காயத்திற்கு எதிராக இது ஒரு பாதுகாப்புப் பாத்திரத்தை வகிக்கிறது. ஃபிமாசார்டன், ரெனின்-ஆஞ்சியோடென்சின் அமைப்பில் செயல்படும் புதிதாக பிரபலமான ஆஞ்சியோடென்சின் II ஏற்பி தடுப்பான், இரத்த அழுத்தத்தைக் குறைக்கிறது; அறுவைசிகிச்சை மூலம் தூண்டப்பட்ட ஒருதலைப்பட்ச சிறுநீர்க்குழாய் அடைப்புடன் எலிகளுக்கு சிகிச்சையளிக்க ஃபிமாசார்டனைப் பயன்படுத்துவது, Nrf2 மற்றும் ஆக்ஸிஜனேற்ற பாதையை அதிகப்படுத்துவதன் மூலம் ஆக்ஸிஜனேற்ற அழுத்தம், வீக்கம் மற்றும் ஃபைப்ரோஸிஸைக் குறைக்கிறது மற்றும் RAS மற்றும் MAPKகளைத் தடுப்பது [129]. சப்பனோன் தென்கிழக்கு ஆசியாவில் பரவலாக விநியோகிக்கப்படுகிறது, அங்கு இது காய்ச்சல் எதிர்ப்பு, ஒவ்வாமை எதிர்ப்பு மற்றும் நரம்புத் தடுப்பு மருந்தாகப் பயன்படுத்தப்படுகிறது; இது Nrf2 ஐ செயல்படுத்துகிறது மற்றும் NF-?B ஐ தடுக்கிறது மற்றும் Nrf2- மற்றும்/அல்லது NF-?B தொடர்பான நோய்களுக்கான சிகிச்சையில் நன்மை பயக்கும் [130]. Bixin orellana விதைகளில் இருந்து பிரித்தெடுக்கப்படும் Bixin மெக்ஸிகோ மற்றும் தென் அமெரிக்காவில் தொற்று மற்றும் அழற்சி நோய்களுக்கு பயன்படுத்தப்படுகிறது; காற்றோட்டத்தால் தூண்டப்பட்ட நுரையீரல் காயத்தைத் தணிக்கவும் சாதாரண நுரையீரல் உருவ அமைப்பை மீட்டெடுக்கவும் இது அழற்சி மத்தியஸ்தர்கள், அல்வியோலர் கேபிலரி கசிவு மற்றும் ஆக்ஸிஜனேற்ற சேதத்தை Nrf2-சார்ந்த முறையில் குறைக்கிறது [131]. எபிகல்லோகேடசின் கேலேட், சல்ஃபோராபேன், ரெஸ்வெராட்ரோல், லைகோபீன் மற்றும் கிரீன் டீ சாறு போன்ற பிற தாவர கலவைகள் Nrf2 சிக்னலிங் பாதை [132], [133], [134] மூலம் அழற்சி நோய்களில் சிகிச்சை விளைவைக் கொண்டுள்ளன. சமீபத்தில் சிட்ரஸ் பழத்தில் உள்ள மற்றொரு பைட்டோகெமிக்கல், எரியோடிக்டியோல், சிஸ்ப்ளேட்டின் தூண்டப்பட்ட சிறுநீரக காயம் மற்றும் செப்சிஸ்-தூண்டப்பட்ட கடுமையான நுரையீரல் காயம் ஆகியவற்றில் அழற்சி எதிர்ப்பு மற்றும் ஆக்ஸிஜனேற்ற விளைவுகளை ஏற்படுத்துவதாக தெரிவிக்கப்பட்டுள்ளது. மேக்ரோபேஜ்களில் சைட்டோகைன்களின் வெளிப்பாடு [2], [135]. இருப்பினும், பல பைட்டோ கெமிக்கல்கள் பல்வேறு மனித நோய்களைத் தடுப்பதற்கும் சிகிச்சையளிப்பதற்கும் பெரும் வாக்குறுதியைக் காட்டுகின்றன, மேலும் சில ஏற்கனவே மருத்துவ பரிசோதனைகள் நிலைக்கு வந்துள்ளன (அட்டவணை 136).
இந்த தாவர கலவைகள் முக்கியமாக கீப்2 இன் சிஸ்டைன் எச்சங்களை மாற்றியமைக்கும் எலக்ட்ரோஃபிலிக் பொருட்களின் வடிவத்தில் Nrf1 சமிக்ஞை பாதையை செயல்படுத்துகிறது, இது ARE உடன் இலவச அணுக்கரு Nrf2 பிணைப்புக்கு வழிவகுக்கிறது, இதன் விளைவாக தொடர்புடைய மரபணுவின் படியெடுத்தல் செயல்படுத்தப்படுகிறது.
புற்றுநோய், இறப்பு, முதுமை, மூளை மற்றும் நடத்தை, இதய நோய் மற்றும் பலவற்றில் சல்போராபேன் மற்றும் அதன் விளைவுகள்
ஐசோதியோசயனேட்டுகள் உங்கள் உணவில் நீங்கள் பெறக்கூடிய மிக முக்கியமான தாவர கலவைகள் ஆகும். இந்த வீடியோவில் நான் அவர்களுக்காக இதுவரை செய்யப்படாத மிக விரிவான வழக்கை உருவாக்குகிறேன். குறுகிய கவனம்? கீழே உள்ள நேரப் புள்ளிகளில் ஒன்றைக் கிளிக் செய்வதன் மூலம் உங்களுக்குப் பிடித்த தலைப்புக்குச் செல்லவும். முழு காலவரிசை கீழே.
முக்கிய பிரிவுகள்:
00:01:14 - புற்றுநோய் மற்றும் இறப்பு
00:19:04 - முதுமை
00:26:30 - மூளை மற்றும் நடத்தை
00:38:06 - இறுதி மறுபரிசீலனை
00:40:27 - டோஸ்
முழு காலவரிசை:
00:00:34 – வீடியோவின் முக்கிய மையமான சல்போராபேன் அறிமுகம்.
00:01:14 - க்ரூசிஃபெரஸ் காய்கறி நுகர்வு மற்றும் அனைத்து காரணங்களின் இறப்பு குறைப்பு.
00:02:12 - புரோஸ்டேட் புற்றுநோய் ஆபத்து.
00:02:23 - சிறுநீர்ப்பை புற்றுநோய் ஆபத்து.
00:02:34 - புகைப்பிடிப்பவர்களுக்கு நுரையீரல் புற்றுநோய் ஆபத்து.
00:02:48 - மார்பக புற்றுநோய் ஆபத்து.
00:03:13 - அனுமானம்: உங்களுக்கு ஏற்கனவே புற்றுநோய் இருந்தால் என்ன செய்வது? (இடையிடல்)
00:03:35 – புற்றுநோய் மற்றும் இறப்பு தொடர்பான தரவுகளை இயக்கும் நம்பத்தகுந்த வழிமுறை.
00:04:38 - சல்போராபேன் மற்றும் புற்றுநோய்.
00:05:32 - எலிகளில் சிறுநீர்ப்பை கட்டி வளர்ச்சியில் ப்ரோக்கோலி முளை சாறு வலுவான விளைவைக் காட்டும் விலங்கு சான்றுகள்.
00:06:06 - புரோஸ்டேட் புற்றுநோயாளிகளில் சல்ஃபோராபேன் நேரடியாக நிரப்புவதன் விளைவு.
00:07:09 - உண்மையான மார்பக திசுக்களில் ஐசோதியோசயனேட் வளர்சிதை மாற்றங்களின் உயிர் குவிப்பு.
00:08:32 - மார்பக புற்றுநோய் ஸ்டெம் செல்களைத் தடுப்பது.
00:08:53 - வரலாற்றுப் பாடம்: பண்டைய ரோமில் கூட பிராசிகாக்கள் ஆரோக்கிய பண்புகளைக் கொண்டிருப்பதாக நிறுவப்பட்டது.
00:09:16 – சல்போராபேன் புற்றுநோய் வெளியேற்றத்தை மேம்படுத்தும் திறன் (பென்சீன், அக்ரோலின்).
00:09:51 - ஆன்டிஆக்ஸிடன்ட் மறுமொழி கூறுகள் வழியாக மரபணு மாற்றமாக NRF2.
00:10:10 – NRF2 ஆக்டிவேஷன் குளுதாதயோன்-எஸ்-கான்ஜுகேட்ஸ் வழியாக புற்றுநோயை வெளியேற்றுவதை எவ்வாறு மேம்படுத்துகிறது.
00:10:34 - பிரஸ்ஸல்ஸ் முளைகள் குளுதாதயோன்-எஸ்-டிரான்ஸ்ஃபெரேஸை அதிகரிக்கின்றன மற்றும் டிஎன்ஏ பாதிப்பைக் குறைக்கின்றன.
00:11:20 - ப்ரோக்கோலி முளை பானம் பென்சீன் வெளியேற்றத்தை 61% அதிகரிக்கிறது.
00:13:31 - ப்ரோக்கோலி ஸ்ப்ரூட் ஹோமோஜெனேட் மேல் சுவாசப்பாதையில் ஆக்ஸிஜனேற்ற நொதிகளை அதிகரிக்கிறது.
00:15:45 - சிலுவை காய்கறி நுகர்வு மற்றும் இதய நோய் இறப்பு.
00:16:55 - ப்ரோக்கோலி ஸ்ப்ரூட் பவுடர் இரத்த கொழுப்பு மற்றும் ஒட்டுமொத்த இதய நோய் அபாயத்தை மேம்படுத்துகிறது வகை 2 நீரிழிவு நோயாளிகள்.
00:19:04 - வயதான பிரிவின் ஆரம்பம்.
00:19:21 - சல்போராபேன்-செறிவூட்டப்பட்ட உணவு வண்டுகளின் ஆயுட்காலம் 15 முதல் 30% வரை அதிகரிக்கிறது (சில நிபந்தனைகளில்).
00:20:34 - நீண்ட ஆயுளுக்கு குறைந்த வீக்கத்தின் முக்கியத்துவம்.
00:22:05 - குரூசிஃபெரஸ் காய்கறிகள் மற்றும் ப்ரோக்கோலி ஸ்ப்ரூட் பவுடர் மனிதர்களில் பல்வேறு வகையான அழற்சி குறிப்பான்களைக் குறைப்பதாகத் தெரிகிறது.
00:23:40 – மிட்-வீடியோ ரீகேப்: புற்றுநோய், வயதான பிரிவுகள்
00:24:14 - முதுமையில் சல்போராபேன் தகவமைப்பு நோயெதிர்ப்புச் செயல்பாட்டை மேம்படுத்தலாம் என்று சுட்டி ஆய்வுகள் தெரிவிக்கின்றன.
00:25:18 – சல்ஃபோராபேன், வழுக்கை மவுஸ் மாதிரியில் முடி வளர்ச்சியை மேம்படுத்தியது. படம் 00:26:10.
00:28:17 - மனச்சோர்வு விவாதத்தின் தொடக்கம் (நம்பத்தகுந்த வழிமுறை மற்றும் ஆய்வுகள்).
00:31:21 – அழுத்தத்தால் தூண்டப்பட்ட மனச்சோர்வின் 10 வெவ்வேறு மாதிரிகளைப் பயன்படுத்தி மவுஸ் ஆய்வு, ஃப்ளூக்ஸெடின் (ப்ரோசாக்) போலவே சல்ஃபோராபேன் பயனுள்ளதாக இருக்கும்.
00:32:00 – எலிகளில் குளுக்கோராபனின் நேரடியாக உட்கொள்வது சமூக தோல்வி மன அழுத்த மாதிரியிலிருந்து மனச்சோர்வைத் தடுப்பதில் இதேபோல் பயனுள்ளதாக இருக்கும் என்று ஆய்வு காட்டுகிறது.
00:33:01 - நியூரோடிஜெனரேஷன் பிரிவின் ஆரம்பம்.
00:33:30 – சல்போராபேன் மற்றும் அல்சைமர் நோய்.
00:33:44 – சல்போராபேன் மற்றும் பார்கின்சன் நோய்.
00:33:51 – சல்போராபேன் மற்றும் ஹங்டிங்டன் நோய்.
00:34:13 - சல்ஃபோராபேன் வெப்ப அதிர்ச்சி புரதங்களை அதிகரிக்கிறது.
00:34:43 - அதிர்ச்சிகரமான மூளை காயம் பிரிவின் ஆரம்பம்.
00:35:55 - சல்ஃபோராபேன் மற்றும் நியூரானல் பிளாஸ்டிசிட்டி.
00:36:32 – எலிகளில் டைப் II நீரிழிவு மாதிரியில் சல்போராபேன் கற்றலை மேம்படுத்துகிறது.
00:37:19 - சல்போராபேன் மற்றும் டுச்சேன் தசைநார் சிதைவு.
00:37:44 - தசை செயற்கைக்கோள் செல்களில் Myostatin தடுப்பு (விட்ரோவில்).
00:38:06 – லேட்-வீடியோ மறுபரிசீலனை: இறப்பு மற்றும் புற்றுநோய், டிஎன்ஏ சேதம், ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் வீக்கம், பென்சீன் வெளியேற்றம், இருதய நோய், வகை II நீரிழிவு, மூளையில் விளைவுகள் (மன அழுத்தம், மன இறுக்கம், ஸ்கிசோஃப்ரினியா, நியூரோடிஜெனரேஷன்), NRF2 பாதை.
00:40:27 - ப்ரோக்கோலி முளைகள் அல்லது சல்ஃபோராபேன் அளவைக் கண்டறிவது பற்றிய எண்ணங்கள்.
00:41:01 - வீட்டில் முளைப்பது பற்றிய நிகழ்வுகள்.
00:43:14 - சமையல் வெப்பநிலை மற்றும் சல்ஃபோராபேன் செயல்பாடு.
00:43:45 - குளுகோராபனினில் இருந்து சல்போராபேன் குடல் பாக்டீரியாவை மாற்றுகிறது.
00:44:24 - காய்கறிகளிலிருந்து செயலில் உள்ள மைரோசினேஸுடன் இணைந்தால் சப்ளிமெண்ட்ஸ் சிறப்பாகச் செயல்படும்.
00:44:56 - சமையல் நுட்பங்கள் மற்றும் சிலுவை காய்கறிகள்.
00:46:06 - ஐசோதியோசயனேட்டுகள் கோயிட்ரோஜன்களாக.
முடிவுகளை
தற்போது, அழற்சியில் Nrf2/Keap1/ARE சிக்னலிங் பாதையின் பங்கில் அதிக ஆராய்ச்சி கவனம் செலுத்தியுள்ளது. Nrf2 ஆல் கட்டுப்படுத்தப்பட்ட நொதிகளில், HO-1 என்பது பிரதிநிதித்துவ அழுத்த பதில் நொதிகளில் ஒன்றாகும். HO-1 முக்கிய அழற்சி எதிர்ப்பு மற்றும் ஆக்ஸிஜனேற்ற பண்புகளைக் கொண்டுள்ளது. பொதுவாக, Nrf2 சமிக்ஞை பாதையானது சைட்டோகைன்கள், கெமோக்கின் வெளியிடும் காரணிகள், MMPகள் மற்றும் பிற அழற்சி மத்தியஸ்தர்களான COX-2 மற்றும் iNOS உற்பத்தியையும் எதிர்மறையாக ஒழுங்குபடுத்துகிறது, இது தொடர்புடைய NF-kB மற்றும் MAPK பாதைகள் மற்றும் வீக்கத்தைக் கட்டுப்படுத்தும் பிற நெட்வொர்க்குகளை நேரடியாகவோ அல்லது மறைமுகமாகவோ பாதிக்கிறது. கீழ்நிலை இலக்கு புரதங்களின் படியெடுத்தல் அல்லது செயல்பாட்டைக் கட்டுப்படுத்த Nrf2 மற்றும் NF-?B சிக்னலிங் பாதைகள் தொடர்பு கொள்கின்றன என்று பரிந்துரைக்கப்படுகிறது. NRf2 மூலம் NF-?B-மத்தியஸ்த டிரான்ஸ்கிரிப்ஷனல் செயல்பாட்டை அடக்குதல் அல்லது செயலிழக்கச் செய்வது பெரும்பாலும் அழற்சியின் ஆரம்ப கட்டத்தில் நிகழ்கிறது, ஏனெனில் NF-?B அழற்சிக்கு சார்பான மத்தியஸ்தர்களின் வரிசையின் டி நோவோ தொகுப்பை ஒழுங்குபடுத்துகிறது. இருப்பினும், Nrf2 மற்றும் JAK/STAT போன்ற பிற சிக்னலிங் பாதைகளுக்கு இடையே இணைப்புகள் உள்ளதா, இயற்கையான தாவர ஆதாரங்களில் இருந்து பெறப்பட்ட தற்போதைய Nrf2 ஆக்டிவேட்டர்களின் முக்கியத்துவம் மற்றும் உயிரியல் செயல்பாட்டை எவ்வாறு மேம்படுத்துவது போன்ற ஆராய்ச்சியில் இன்னும் சில வரம்புகள் உள்ளன. மேலும் இந்த சேர்மங்களின் இலக்கை அதிகரிக்கவும். இவற்றுக்கு மேலும் சோதனை சரிபார்ப்பு தேவைப்படுகிறது.
கூடுதலாக, Nrf2 சிக்னலிங் பாதையானது > 600 மரபணுக்களைக் கட்டுப்படுத்தலாம் [163], இதில் > 200 சைட்டோபுரோடெக்டிவ் புரதங்களைக் குறியாக்குகிறது [164] அவை வீக்கம், புற்றுநோய், நரம்பியக்கடத்தல் நோய்கள் மற்றும் பிற முக்கிய நோய்களுடன் தொடர்புடையவை [165]. Nrf2 சிக்னலிங் பாதை பல புற்றுநோய்களில் ஒழுங்குபடுத்தப்படவில்லை, இதன் விளைவாக மாறுபாடு வெளிப்பாடு Nrf2 சார்ந்த மரபணு பேட்டரி என்று பரிந்துரைக்கும் வளர்ந்து வரும் சான்றுகள். மேலும், ஆக்ஸிஜனேற்ற அழுத்தம் தொடர்பான நோய்களில் குறிப்பாக புற்றுநோயில் வீக்கம் முக்கிய பங்கு வகிக்கிறது. அழற்சியை எதிர்ப்பதற்கு பல Nrf2 ஆக்டிவேட்டர்களின் பயன்பாடு Nrf2 கீழ்நிலை மரபணுக்களின் மாறுபட்ட வெளிப்பாட்டை ஏற்படுத்தலாம், இது புற்றுநோயியல் மற்றும் கீமோ மற்றும்/அல்லது ரேடியோ சிகிச்சைக்கு எதிர்ப்பைத் தூண்டுகிறது. எனவே, அதன் ப்ளியோட்ரோபிக் விளைவுகளைக் குறைக்க Nrf2 இன் மிகவும் குறிப்பிட்ட ஆக்டிவேட்டர்கள் உருவாக்கப்படலாம். Nrf2 இன் பல ஆக்டிவேட்டர்கள் ஆக்ஸிஜனேற்ற அழுத்தம் தொடர்பான நோய்களில் அழற்சி எதிர்ப்பு செயல்பாடுகளில் குறிப்பிடத்தக்க முன்னேற்றத்தைக் காட்டியுள்ளன. FDA ஆல் அங்கீகரிக்கப்பட்ட Nrf2 ஆக்டிவேட்டரின் சிறந்த உதாரணம் மற்றும் மல்டிபிள் ஸ்களீரோசிஸ் (MS) போன்ற அழற்சி நோய்களுக்கான சிகிச்சையில் பரவலாகப் பயன்படுத்தப்படுவது டைமெத்தில் ஃபுமரேட் ஆகும். டெக்ஃபிடெரா (பயோஜென் மூலம் டைமெத்தில் ஃபுமரேட்டின் பதிவு செய்யப்பட்ட பெயர்) அதிக எண்ணிக்கையிலான நோயாளிகளில் மல்டிபிள் ஸ்களீரோசிஸின் மறுபிறப்பு வடிவங்களுக்கு சிகிச்சையளிக்க திறம்பட பயன்படுத்தப்படுகிறது [152]. இருப்பினும், அழற்சி நோய்களுக்கு சிகிச்சையளிக்க Nrf2 ஆக்டிவேட்டர்களைப் பயன்படுத்துவதன் செயல்திறனுக்கு Nrf2 இன் தீங்கு விளைவிக்கும் விளைவுகளைத் தவிர்க்க மேலும் சரிபார்ப்பு தேவைப்படுகிறது. எனவே, Nrf2 ஆல் மத்தியஸ்தம் செய்யப்பட்ட அழற்சி எதிர்ப்பு நடவடிக்கைக்கான சிகிச்சையின் வளர்ச்சி குறிப்பிடத்தக்க மருத்துவ தாக்கத்தை ஏற்படுத்தக்கூடும். உலகெங்கிலும் உள்ள Nrf2 சமிக்ஞை பாதையின் தொடர்ச்சியான ஆய்வுகள் வீக்கத்தின் அறிகுறிகளைக் கட்டுப்படுத்தவும், புற்றுநோய் மற்றும் நரம்பியக்கடத்தல் மற்றும் பிற முக்கிய நோய்களைத் தடுப்பதற்கும் சிகிச்சையளிப்பதற்கும் அதிக இலக்கு கொண்ட சிகிச்சை முகவர்களை உருவாக்குவதற்கு அர்ப்பணித்துள்ளன.
முடிவில், Nrf2 மனித உடலில் ஆக்ஸிஜனேற்ற அழுத்தத்தின் அளவை உணர்கிறது மற்றும் இறுதியில் ஆக்ஸிஜனேற்ற மற்றும் நச்சுத்தன்மையற்ற நொதிகள் மற்றும் மரபணுக்களை ஒழுங்குபடுத்த உதவுகிறது. அதிகரித்த அளவு ஆக்ஸிஜனேற்ற அழுத்தத்தால் ஏற்படும் நாள்பட்ட அழற்சி நரம்பியக்கடத்தல் நோய்களுடன் தொடர்புடையதாக இருப்பதால், Nrf2 ஒரு முக்கிய பாத்திரத்தை வகிக்க முடியும் அல்சைமர் நோய் போன்ற உடல்நலப் பிரச்சினைகளின் சிகிச்சையில், மற்றவற்றுடன். எங்கள் தகவலின் நோக்கம் உடலியக்க மற்றும் முதுகெலும்பு சுகாதார பிரச்சினைகள் மட்டுமே. விஷயத்தைப் பற்றி விவாதிக்க, தயவு செய்து டாக்டர் ஜிமினெஸைக் கேட்கவும் அல்லது எங்களை இங்கே தொடர்பு கொள்ளவும்915-850-0900.
கூடுதல் தலைப்பு விவாதம்: அறுவை சிகிச்சை இல்லாமல் முழங்கால் வலியை நீக்குதல்
முழங்கால் வலி என்பது நன்கு அறியப்பட்ட அறிகுறியாகும், இது பல்வேறு முழங்கால் காயங்கள் மற்றும்/அல்லது நிலைமைகள் காரணமாக ஏற்படலாம்.விளையாட்டு காயங்கள். முழங்கால் மனித உடலில் மிகவும் சிக்கலான மூட்டுகளில் ஒன்றாகும், ஏனெனில் இது நான்கு எலும்புகள், நான்கு தசைநார்கள், பல்வேறு தசைநாண்கள், இரண்டு மெனிசிஸ் மற்றும் குருத்தெலும்பு ஆகியவற்றின் குறுக்குவெட்டுகளால் ஆனது. அமெரிக்கன் அகாடமி ஆஃப் ஃபேமிலி பிசிஷியன்களின் கூற்றுப்படி, முழங்கால் வலிக்கான பொதுவான காரணங்களில் பட்டேலர் சப்லக்சேஷன், பட்டேலர் டெண்டினிடிஸ் அல்லது ஜம்பர்ஸ் முழங்கால் மற்றும் ஓஸ்குட்-ஸ்க்லாட்டர் நோய் ஆகியவை அடங்கும். முழங்கால் வலி 60 வயதுக்கு மேற்பட்டவர்களுக்கு ஏற்பட வாய்ப்புகள் இருந்தாலும், குழந்தைகள் மற்றும் இளம்பருவத்தினருக்கும் முழங்கால் வலி ஏற்படலாம். முழங்கால் வலியை RICE முறைகளைப் பின்பற்றி வீட்டிலேயே சிகிச்சை செய்யலாம், இருப்பினும், கடுமையான முழங்கால் காயங்களுக்கு உடலியக்க சிகிச்சை உட்பட உடனடி மருத்துவ கவனிப்பு தேவைப்படலாம். �
அல்சைமர் நோய் மற்றும் பார்கின்சன் நோய் போன்ற நியூரோடிஜெனரேட்டிவ் நோய்கள் உலகளவில் மில்லியன் கணக்கான நபர்களை பாதிக்கின்றன. பல நரம்பியக்கடத்தல் நோய்களின் அறிகுறிகளுக்கு சிகிச்சையளிக்க பல்வேறு சிகிச்சை விருப்பங்கள் உள்ளன, இருப்பினும் முடிவுகள் பெரும்பாலும் குறைவாகவே இருக்கும். உள் மற்றும் வெளிப்புற காரணிகளால் ஏற்படும் ஆக்ஸிஜனேற்ற அழுத்தம் நரம்பியக்கடத்தல் நோய்களின் வளர்ச்சிக்கு ஒரு காரணமாக இருக்கலாம் என்று ஆராய்ச்சி ஆய்வுகள் கண்டறிந்துள்ளன. தி டிரான்ஸ்கிரிப்ஷன் காரணி, Nrf2, ஆக்ஸிஜனேற்ற அழுத்தத்திற்கு எதிராக ஒரு முக்கிய பாதுகாப்பு பொறிமுறையாக செயல்பட தீர்மானிக்கப்பட்டுள்ளது. அதன் விளைவுகளைக் காண்பிப்பதே கீழே உள்ள கட்டுரையின் நோக்கம் நியூரோடிஜெனரேட்டிவ் நோய்களில் Nrf2.
டிரான்ஸ்கிரிப்ஷன் காரணி NRF2 மூலம் புரோட்டியோஸ்டாசிஸின் பண்பேற்றம்
நியூரோடிஜெனரேட்டிவ் நோய்கள் குறிப்பிட்ட புரதத் தொகுப்புகளின் திரட்சியுடன் இணைக்கப்பட்டுள்ளன, இது காயமடைந்த மூளைக்கும் புரோட்டியோஸ்டாசிஸ் இழப்புக்கும் இடையே ஒரு நெருக்கமான தொடர்பைக் குறிக்கிறது. சிக்னலிங் பாதைகள், மரபணு வெளிப்பாடு மற்றும் புரதச் சிதைவு அமைப்புகளின் ஒழுங்குமுறையை ஒருங்கிணைக்கும் பரந்த நெட்வொர்க்கிற்கு நன்றி, புரோட்டியோமின் மிகுதி மற்றும் மடிப்புகளை செல்கள் கட்டுப்படுத்தும் அனைத்து செயல்முறைகளையும் புரோட்டியோஸ்டாஸிஸ் குறிக்கிறது. டிரான்ஸ்கிரிப்ஷன் காரணி NRF2 (அணு காரணி (எரித்ராய்டு-பெறப்பட்ட 2) போன்ற 2) மூலம் செலுத்தப்படும் புரோட்டியோஸ்டாசிஸின் டிரான்ஸ்கிரிப்ஷனல் பண்பேற்றம் பற்றிய மிகவும் பொருத்தமான கண்டுபிடிப்புகளை சுருக்கமாகக் கூற இந்த மதிப்பாய்வு முயற்சிக்கிறது. NRF2 ஆனது ஆன்டிஆக்ஸிடன்ட் செல் பதிலின் முதன்மை சீராக்கியாக பாரம்பரியமாக கருதப்படுகிறது, இருப்பினும் இது தற்போது புரோட்டியோஸ்டாசிஸை பராமரிக்க கடத்தும் இயந்திரத்தின் முக்கிய அங்கமாக உருவாகி வருகிறது. நாம் விவாதிப்பது போல, NRF2 ஆனது ஒரு ஒருங்கிணைந்த மற்றும் நீடித்த டிரான்ஸ்கிரிப்ஷனல் பதிலை உருவாக்குவதற்காக தவறாக மடிக்கப்பட்ட புரத திரட்சியிலிருந்து பெறப்பட்ட அவசரகால சமிக்ஞைகளை தொகுக்கும் மையமாக கருதப்படலாம். எண்டோபிளாஸ்மிக் ரெட்டிகுலம் உடலியல், புரோட்டீசோம் மற்றும் ஆட்டோபேஜி ஆகியவற்றின் பராமரிப்பில் ஈடுபட்டுள்ள மரபணுக்களின் கட்டுப்பாடு தொடர்பான NRF2 இன் செயல்பாடுகளால் இது அடையப்படுகிறது.
முக்கிய வார்த்தைகள்:நியூரோடிஜெனரேட்டிவ் நோய்கள், விரிவடைந்த புரத பதில், புரோட்டீசோம், யுபிக்விடின், தன்னியக்கவியல், ஆக்ஸிஜனேற்ற அழுத்தம்
நியூக்ளியர் ஃபேக்டர் (எரித்ராய்டு-பெறப்பட்ட 2)-போன்ற 2 (NRF2) என்பது செல்லுலார் ஹோமியோஸ்டாசிஸின் முதன்மை சீராக்கியாக தற்போது கருதப்படுகிறது. இது 250 க்கும் மேற்பட்ட மரபணுக்களின் அடிப்படை மற்றும் அழுத்த-தூண்டக்கூடிய வெளிப்பாட்டைக் கட்டுப்படுத்துகிறது, அவை பொதுவாக சிஸ்-ஆக்டிங் மேம்பாட்டாளரைப் பகிர்ந்து கொள்கின்றன, அவை ஆக்ஸிஜனேற்ற மறுமொழி உறுப்பு (ARE) [1], [2], [3], [4], [5]. இந்த மரபணுக்கள் கட்டம் I, II மற்றும் III நச்சுத்தன்மை வினைகள், குளுதாதயோன் மற்றும் பெராக்ஸிரெடாக்சின்/தியோரெடாக்சின் வளர்சிதை மாற்றம், பென்டோஸ் பாஸ்பேட் பாதை மற்றும் மாலிக் என்சைம் மூலம் NADPH உற்பத்தி, கொழுப்பு அமில ஆக்சிஜனேற்றம், இரும்பு வளர்சிதை மாற்றம் மற்றும் புரோட்டியோஸ்டாஸிஸ் [6] ஆகியவற்றில் பங்கேற்கின்றன. இந்த பரந்த சைட்டோபுரோடெக்டிவ் செயல்பாடுகளின் அடிப்படையில், NRF2 இல் உள்ள ஒரு மருந்தியல் வெற்றியானது ஆக்ஸிஜனேற்ற, அழற்சி மற்றும் புரோட்டியோடாக்ஸிக் அழுத்தம் உள்ளிட்ட நாள்பட்ட நோய்களின் முக்கிய குற்றவாளிகளின் விளைவைத் தணிக்கக்கூடும். ஆக்ஸிஜனேற்ற பாதுகாப்பின் பண்பேற்றத்தில் NRF2 இன் பங்கு மற்றும் அழற்சியின் தீர்மானம் பல ஆய்வுகளில் குறிப்பிடப்பட்டுள்ளது ([7] இல் மதிப்பாய்வு செய்யப்பட்டது). இங்கே, புரோட்டியோஸ்டாசிஸில் அதன் பங்கில் கவனம் செலுத்துவோம், அதாவது புரத தொகுப்பு, மடிப்பு, கடத்தல் மற்றும் சீரழிவு ஆகியவற்றின் ஹோமியோஸ்ட்டிக் கட்டுப்பாடு. நியூரோடிஜெனரேட்டிவ் நோய்களின் பின்னணியில் எடுத்துக்காட்டுகள் வழங்கப்படும்.
நியூரோடிஜெனரேட்டிவ் நோய்களில் NRF2 செயல்பாட்டைப் பாதிக்கும் புரோட்டியோஸ்டாசிஸ் இழப்பு
நியூரோடிஜெனரேடிவ் நோய்களின் பொதுவான அடையாளம் சில புரதங்களின் மாறுபட்ட திரட்டல் நிகழ்வாகும். இவ்வாறு, பார்கின்சன் நோய் (PD), ?-அமிலாய்டு (A?) பிளேக்குகள் மற்றும் அல்சைமர் நோய் (AD), huntingtin (Htt) இல் உள்ள ஹைப்பர்-பாஸ்போரிலேட்டட் TAU நியூரோபிப்ரில்லரி சிக்கல்களில் ?-synuclein (?-SYN) இன் தவறாக மடிக்கப்பட்ட புரதத் திரட்டுகள் காணப்படுகின்றன. ஹண்டிங்டன் நோய் (HD), சூப்பர் ஆக்சைடு டிஸ்முடேஸ் 1 (SOD1) மற்றும் அமியோட்ரோபிக் லேட்டரல் ஸ்களீரோசிஸ் (ALS) இல் TAR DNA பிணைப்பு புரதம் 43 (TDP-43), ஸ்பாங்கிஃபார்ம் என்செபலோபதியில் உள்ள ப்ரியான் புரதம் (PrP) போன்றவை. புரதத் திரட்டுகள் பலவற்றில் தாக்கத்தை ஏற்படுத்தலாம். செல்லுலார் பாதைகள், இது NRF2 அளவுகள் மற்றும் செயல்பாட்டை பாதிக்கலாம்.
NRF2 செயல்பாட்டைக் கட்டுப்படுத்தும் பல்வேறு அடுக்குகள்
உடலியல் நிலைமைகளின் கீழ், செல்கள் அதன் விரைவான விற்றுமுதல் காரணமாக குறைந்த NRF2 புரத அளவுகளை வெளிப்படுத்துகின்றன. வெவ்வேறு தூண்டுதல்களுக்கு பதிலளிக்கும் விதமாக, NRF2 புரதம் குவிந்து, கருவுக்குள் நுழைந்து ARE-கொண்ட மரபணுக்களின் படியெடுத்தலை அதிகரிக்கிறது. எனவே, NRF2 புரத அளவுகளின் மேலாண்மை நேர்மறை மற்றும் எதிர்மறை உள்ளீட்டு சமிக்ஞைகளை ஒருங்கிணைக்க வேண்டிய ஒரு முக்கிய புள்ளியாகும். நாம் மேலும் விவாதிப்பதைப் போல, விரைவான மற்றும் திறமையான பதிலைத் திட்டமிடுவதற்கு NRF2 பல்வேறு ஒன்றுடன் ஒன்று இயங்கும் வழிமுறைகளால் செயல்படுத்தப்படுகிறது, ஆனால் மறுபுறம் NRF2 அதன் பதிலை அணைக்க இரண்டாவது கட்டத்தில் தடுக்கப்படலாம்.
உன்னதமான பார்வையில், ஆக்ஸிஜனேற்ற அல்லது எலக்ட்ரோஃபிலிக் கலவைகளுக்கு செல்லுலார் பதிலின் விளைவாக NRF2 செயல்படுத்தப்படுகிறது. இது சம்பந்தமாக, ubiquitin E3 லிகேஸ் அடாப்டர் கெல்ச் போன்ற ECH-தொடர்புடைய புரதம் 1 (KEAP1) ஒரு முக்கிய பங்கு வகிக்கிறது. மூலக்கூறு விவரங்கள் பிரிவு 4.1 இல் மேலும் குறிப்பிடப்படும். சுருக்கமாக, NRF1 எங்கும் பரவுதல் மற்றும் புரோட்டீசோமால் சிதைவுக்கு வழிவகுக்கும் முக்கியமான சிஸ்டைன் எச்சங்கள் காரணமாக KEAP2 ஒரு ரெடாக்ஸ் சென்சாராக செயல்படுகிறது. இந்த உன்னதமான பண்பேற்றத்துடன் கூடுதலாக, NRF2 நிகழ்வுகளை சமிக்ஞை செய்வதன் மூலம் ஆழமாக கட்டுப்படுத்தப்படுகிறது. உண்மையில், வெவ்வேறு கைனேஸ்கள் பாஸ்போரிலேட் மற்றும் NRF2 ஐ ஒழுங்குபடுத்துகின்றன. உதாரணமாக, NRF2 ஐ மைட்டோஜென் ஆக்டிவேட்டட் புரோட்டீன் கைனேஸ்கள் (MAPKs) மூலம் பாஸ்போரிலேட் செய்ய முடியும், இருப்பினும் NRF2 செயல்பாட்டில் அதன் பங்களிப்பு தெளிவாக இல்லை [8], [9], [10], [11]. PKA கைனேஸ் மற்றும் சில PKC ஐசோசைம்கள் [12], CK2 [13] அல்லது Fyn [14] பாஸ்போரிலேட் NRF2 அதன் நிலைத்தன்மையை மாற்றியமைக்கிறது. எங்கள் குழுவின் முந்தைய வேலை கிளைகோஜன் சின்தேஸ் கின்ஸ்-3? (GSK-3?) NRF2 ஐ அணுக்கரு விலக்கு மற்றும் புரோட்டீசோமால் சிதைவு [15], [25], [26], [27], [28], [29], [30] மூலம் தடுக்கிறது. மூலக்கூறு விவரங்கள் பிரிவு 4.1 இல் விவாதிக்கப்படும். மேலும், NRF2 மற்ற வகை ஒழுங்குமுறைகளுக்கு சமர்ப்பிக்கப்படுகிறது. உதாரணமாக, CBP/p2 மூலம் NRF300 அசிடைலேஷன் அதன் செயல்பாட்டை அதிகரிக்கிறது [17], அது miR153, miR27a, miR142-5p, மற்றும் miR144 [16] அல்லது சைட்டோசின்-குவானைன் (CG) தீவுகளை ஊக்குவிக்கும் மெத்திலேஷன் மூலம் தடுக்கப்படுகிறது. [2].
NRF2 ஒழுங்குமுறை வழிமுறைகளில் புரதத் திரட்டுகளின் தாக்கம்
இந்த பிரிவில், தவறான மடிந்த புரதத்தின் குவிப்பு NRF2 செயல்பாட்டை எவ்வாறு பாதிக்கும் என்பதில் கவனம் செலுத்துவோம், இது மேலே குறிப்பிட்டுள்ள சில பாதைகளை விளக்க எடுத்துக்காட்டுகளாக வழங்குகிறது. முதலாவதாக, புரதக் குவிப்பு ஆக்ஸிஜனேற்ற சேதத்துடன் இறுக்கமாக இணைக்கப்பட்டுள்ளது என்பதை நாம் கருத்தில் கொள்ள வேண்டும். உண்மையில், தவறாக மடிக்கப்பட்ட புரதக் குவிப்பு மற்றும் திரட்டுதல் ஆகியவை மைட்டோகாண்ட்ரியா மற்றும் பிற மூலங்களிலிருந்து எதிர்வினை ஆக்ஸிஜன் இனங்களின் (ROS) அசாதாரண உற்பத்தியைத் தூண்டுகின்றன [19]. மேலே குறிப்பிட்டுள்ளபடி, ROS ஆனது KEAP1 இன் ரெடாக்ஸ்-சென்சிட்டிவ் சிஸ்டைன்களை மாற்றியமைக்கும், இது NRF2 இன் வெளியீடு, உறுதிப்படுத்தல் மற்றும் அணுக்கரு பரவலுக்கு வழிவகுக்கும்.
புரோட்டினோபதிகளைப் பொறுத்தவரை, NRF2 ஐப் பாதிக்கக்கூடிய ஒழுங்குபடுத்தப்படாத சிக்னலிங் நிகழ்வுகளின் உதாரணம் GSK-3 இன் ஹைபராக்டிவேஷன் மூலம் வழங்கப்படுகிறது? கி.பி. TAU கைனேஸ் என்றும் அழைக்கப்படும் GSK-3?, இந்த நுண்குழாய்-தொடர்புடைய புரதத்தின் பாஸ்போரிலேஷனில் பங்கேற்கிறது, இதன் விளைவாக அதன் ஒருங்கிணைப்பு, நியூரோபிப்ரில்லரி சிக்கல்கள் மற்றும் அச்சுப் போக்குவரத்தில் தடங்கல் ஏற்படுகிறது ([20] இல் மதிப்பாய்வு செய்யப்பட்டது). மறுபுறம், GSK-3? மேலே குறிப்பிட்டுள்ளபடி NRF2 அளவுகளையும் செயல்பாட்டையும் வியத்தகு முறையில் குறைக்கிறது. பரவலாக ஏற்றுக்கொள்ளப்படாவிட்டாலும், அமிலாய்டு அடுக்கானது அந்த நச்சுத்தன்மையான A? ஒலிகோமர்கள் GSK-3 ஐ அதிகரிக்குமா? TAU ஹைப்பர்-பாஸ்போரிலேஷன் மற்றும் நியூரானின் இறப்பு [21], [22] ஆகியவற்றுடன் இணைந்து செயல்படும். A எப்படி என்பதை விளக்க பல்வேறு மாதிரிகள் உள்ளன? GSK3-க்கு சாதகமா? செயல்பாடு. உதாரணமாக, ஏ? இன்சுலின் ஏற்பியுடன் பிணைக்கிறது மற்றும் PI3K மற்றும் AKT சிக்னலிங் பாதைகளைத் தடுக்கிறது, அவை GSK-3 ஐப் பராமரிக்க முக்கியமானதா? அதன் N-டெர்மினல் Ser9 எச்சத்தில் பாஸ்போரிலேஷன் மூலம் செயலிழக்கப்பட்டது [23]. மறுபுறம், எக்ஸ்ட்ராசெல்லுலர் ஏ? Frizzled receptors உடன் தொடர்பு கொள்கிறது, WNT சிக்னலைத் தடுக்கிறது [24] மற்றும் மீண்டும் செயலில் உள்ள GSK-3 வெளியீடு?. சுருக்கமாக, ஏ? திரட்சியானது GSK-3 இன் அசாதாரண அதிவேக செயலுக்கு வழிவகுக்கிறது, இதனால் பொருத்தமான NRF2 பதிலை பாதிக்கிறது.
பின்வரும் பிரிவில் விவாதிக்கப்பட்டபடி, தவறாக மடிக்கப்பட்ட புரதங்கள் PERK மற்றும் MAPKகளை செயல்படுத்த வழிவகுக்கும், இது NRF2 [31], [8], [9], [10], [11] ஆகியவற்றைக் கட்டுப்படுத்துகிறது. மேலும், ஒழுங்குபடுத்தப்படாத CBP/p300 செயல்பாடு பல புரோட்டினோபதிகளில் பதிவாகியுள்ளது [32] மேலும் AD மூளையில் DNA மெத்திலேஷனில் உலகளாவிய குறைவு காட்டப்பட்டது [33], எனவே NRF2 ஒழுங்குமுறையில் இந்த கண்டுபிடிப்புகளின் பொருத்தத்தை ஆராய்வதற்கான ஆதாரங்களை வழங்குகிறது.
PD மற்றும் AD நோயாளிகளின் நெக்ரோப்ஸிகளில் NRF2 புரத அளவுகள் மற்றும் ஹீம் ஆக்சிஜனேஸ் 1 (HMOX1), NADPH குயினோன் ஆக்சிடேஸ் 1 (NQO1), p62, போன்ற சில இலக்குகளில் அதிகரிப்பதை நாங்களும் மற்றவர்களும் அவதானித்துள்ளோம். இம்யூனோஹிஸ்டோ கெமிஸ்ட்ரி மூலம் [34], [35], [36], [37], [38], [39]. இந்த நோய்களில் NRF2 இன் மேல்-ஒழுங்குமுறையானது ஹோமியோஸ்ட்டிக் மதிப்புகளை மீட்டெடுக்க நோயுற்ற மூளையின் தோல்வியுற்ற முயற்சியாக விளக்கப்படுகிறது. இருப்பினும், மற்றொரு ஆய்வு NRF2 முக்கியமாக AD ஹிப்போகாம்பல் நியூரான்களின் சைட்டோபிளாஸில் உள்ளமைக்கப்பட்டுள்ளது, இது மூளையில் NRF2 டிரான்ஸ்கிரிப்ஷனல் செயல்பாட்டைக் குறைக்கிறது [40]. இந்த அவதானிப்புகளின் ஏற்றத்தாழ்வு நியூரோடிஜெனரேஷனின் முற்போக்கான நிலைகளில் NRF2 ஐக் கட்டுப்படுத்தும் காரணிகளில் ஏற்படும் மாற்றங்களுடன் தொடர்புடையது என்பது சிந்திக்கத்தக்கது.
மூன்று முக்கிய அமைப்புகள் புரோட்டியோஸ்டாசிஸுக்கு பங்களிக்கின்றன, அதாவது விரிந்த புரத பதில் (UPR), எபிக்விடின் புரோட்டீசோம் அமைப்பு (UPS) மற்றும் தன்னியக்கவியல். அடுத்து, புரோட்டீன் வழித்தோன்றல் இயந்திரங்களுடன் புரதத் திரட்டுகளால் செயல்படுத்தப்படும் அவசரகால சமிக்ஞைகளை இணைக்கும் மையமாக NRF2 ஐக் கற்பனை செய்வதற்கான ஆதாரங்களை முன்வைக்கிறோம்.
ER இல் உள்ள ஆக்ஸிஜனேற்ற புரத மடிப்பு பல வேறுபட்ட பாதைகளால் இயக்கப்படுகிறது, இதில் மிகவும் பாதுகாக்கப்பட்ட புரதம் டைசல்பைட்-ஐசோமரேஸ் (PDI) மற்றும் சல்பைட்ரைல் ஆக்சிடேஸ் எண்டோபிளாஸ்மிக் ஆக்சிடோரேடக்டின் 1 (ERO1? மற்றும் ERO1? பாலூட்டிகளில்) டைசல்பைடு நன்கொடையாளர். சுருக்கமாக, PDI ஆனது அதன் சொந்த சிஸ்டைன் அமினோ அமிலங்களின் குறைப்பு மற்றும் ஆக்சிஜனேற்றம் காரணமாக, புரதங்களுக்குள் உள்ள சிஸ்டைன் எச்சங்களுக்கிடையில் டிஸல்பைடு பிணைப்புகளை உருவாக்குதல் மற்றும் உடைத்தல் ஆகியவற்றை ஊக்குவிக்கிறது. PDI ஆனது ஹவுஸ் கீப்பிங் என்சைம் ERO1 இன் செயல்பாட்டின் மூலம் மறுசுழற்சி செய்யப்படுகிறது, இது டிசல்பைட் பிணைப்புகளை PDI இல் மீண்டும் அறிமுகப்படுத்துகிறது [41]. மூலக்கூறு ஆக்ஸிஜன் என்பது ERO1 இன் முனைய எலக்ட்ரான் ஏற்பியாகும், இது உற்பத்தி செய்யப்படும் ஒவ்வொரு டைசல்பைட் பிணைப்பிற்கும் ஹைட்ரஜன் பெராக்சைடை ஸ்டோச்சியோமெட்ரிக் அளவு உருவாக்குகிறது [42]. பெராக்ஸிடேஸ்கள் (PRX4) மற்றும் குளுதாதயோன் பெராக்ஸிடேஸ்கள் (GPX7 மற்றும் GPX8) ஆகியவை ER இல் ஹைட்ரஜன் பெராக்சைடைக் குறைப்பதற்கான முக்கிய நொதிகளாகும். இந்த ஆக்சிடோ-குறைப்பு அமைப்பு சரியாக வேலை செய்யாதபோது, ER இல் தவறாக மடிக்கப்பட்ட புரதங்களின் திரட்சி ஏற்படுகிறது மற்றும் ER ஹோமியோஸ்டாசிஸை மீண்டும் நிலைநிறுத்த சைட்டோபிளாசம் மற்றும் நியூக்ளியஸுக்கு மடிக்கப்பட்ட புரத மறுமொழி (UPR) என்ற சமிக்ஞைகளின் தொகுப்பு அனுப்பப்படுகிறது [43]. யூகாரியோட்களில் ER அழுத்தத்தை உணர மூன்று சவ்வு-தொடர்புடைய புரதங்கள் அடையாளம் காணப்பட்டுள்ளன: டிரான்ஸ்கிரிப்ஷன் காரணி 6 (ATF6), கணைய ER eIF2 ஐ செயல்படுத்துவது? கைனேஸ் (PERK, இரட்டை இழையுடைய ஆர்என்ஏ-செயல்படுத்தப்பட்ட புரதம் கைனேஸ் போன்ற ER கைனேஸ்), மற்றும் இனோசிட்டால்-தேவையான கைனேஸ்1 (IRE1). ஒவ்வொரு சென்சாரின் லுமினல் டொமைனும் குளுக்கோஸ்-ஒழுங்குபடுத்தப்பட்ட புரதம் (GRP78/BIP) எனப்படும் 78 kDa சேப்பரோனுடன் பிணைக்கப்பட்டுள்ளது. மடிந்த புரதங்களை பிணைக்க ER அழுத்தத்தின் மீது BIP பிரிகிறது, இது மூன்று சென்சார்களை செயல்படுத்துவதற்கு வழிவகுக்கிறது [44].
NRF2 மற்றும் அதன் ஹோமோலாக் NRF1, ஆக்ஸிஜனேற்ற மறுமொழியுடன் தொடர்புடையது, UPR ஐ அணுக்கருவுக்கு கடத்துவதில் பங்கேற்கிறது. NRF1 ஐப் பொறுத்தவரை, இந்த புரதம் ER சவ்வில் அமைந்துள்ளது மற்றும் டிக்ளைகோசைலேஷன் அல்லது பிளவுகளின் மீது அணுக்கரு இடமாற்றத்திற்கு உட்படுகிறது. பின்னர், UPR செயல்படுத்தல் NRF1 இன் செயலாக்கத்திற்கும், அணுசக்தி பெட்டியில் விளைந்த துண்டின் அணுசக்தி குவிப்புக்கும் வழிவகுக்கிறது. இருப்பினும், இந்த NRF1 துண்டின் ARE-கொண்ட மரபணுக்களை மாற்றும் திறன் இன்னும் விவாதத்தில் உள்ளது [45].
Glover-Cutter மற்றும் சக பணியாளர்கள் C. elegans, SKN-2 இன் NRF1 ஆர்த்தோலாக்கை பல்வேறு ER அழுத்தங்களுடன் செயல்படுத்துவதைக் காட்டினர். அதிகரித்த SKN-1 வெளிப்பாடு IRE1 அல்லது PERK worm orthologues [46] உட்பட பல்வேறு UPR மத்தியஸ்தர்களைச் சார்ந்தது. PERK-குறைபாடுள்ள உயிரணுக்களில், குறைபாடுள்ள புரதச் சேர்க்கையானது எண்டோஜெனஸ் பெராக்சைடுகளின் திரட்சிக்கு வழிவகுக்கிறது மற்றும் அதைத் தொடர்ந்து அப்போப்டொசிஸ் [47]. இந்த பெராக்சைடுகளிலிருந்து ER ஐப் பாதுகாக்க PERK பயன்படுத்தும் எஃபெக்டர் NRF2 ஆக இருக்கலாம், ஏனெனில் Ser2 இல் PERK பாஸ்போரிலேட்டுகள் NRF40 என்று தெரிவிக்கப்பட்டுள்ளது, இதனால் அதன் சிதைவை KEAP1 தடுக்கிறது [31]. IRE1 [2] இன் TRAF1-மத்தியஸ்த கைனேஸ் நடவடிக்கை மூலம் ASK48 இன் தூண்டல் இந்த வழியில் ஒரு பங்கைக் கொண்டிருக்கக்கூடும். NRF2 ஐ ஒழுங்குபடுத்துவதில் MAPKகளின் பங்கு இன்னும் சர்ச்சைக்குரியதாக இருந்தாலும், IRE1-TRAF2-ASK1-JNK பாதை NRF2 [49] (படம் 1) செயல்படுத்தலாம் என்று சமீபத்தில் பரிந்துரைக்கப்பட்டது. சுவாரஸ்யமாக, C. elegans மற்றும் மனித உயிரணுக்களில், IRE1 கைனேஸின் சிஸ்டைன் சல்பெனிலேஷன் அதன் செயல்படுத்தும் வளையத்தில் IRE1-மத்தியஸ்தம் UPR ஐத் தடுக்கிறது மற்றும் NRF38 ஆல் இயக்கப்படும் p2 ஆக்ஸிஜனேற்ற பதிலைத் தொடங்குகிறது என்று புதிய சான்றுகள் தெரிவிக்கின்றன. P1 மற்றும் NRF38 [2] ஆகியவற்றைச் செயல்படுத்தும் சைட்டோபிளாஸ்மிக் சென்டினலாக IRE50 ஒரு பண்டைய செயல்பாட்டைக் கொண்டுள்ளது என்று தரவு தெரிவிக்கிறது.
படம் 1 UPR மூலம் NRF2 ஒழுங்குமுறை. எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தின் உள்ளே விரிந்த அல்லது தவறாக மடிக்கப்பட்ட புரதங்களின் குவிப்பு விரிந்த புரத மறுமொழியை (UPR) தொடங்கலாம். முதலாவதாக, மடிக்கப்பட்ட/தவறாக மடிக்கப்பட்ட புரதங்களை பிணைக்க, ER சென்சார்கள் IRE1 மற்றும் PERK இன் இன்ட்ராலூமினல் டொமைனில் இருந்து சாப்பரோன் BIP வெளியிடப்படுகிறது. இது அவர்களின் சைட்டோசோலிக் டொமைன்களின் டைமரைசேஷன் மற்றும் டிரான்ஸ்-ஆட்டோ-பாஸ்போரிலேஷன் ஆகியவற்றை செயல்படுத்துகிறது. PERK செயல்படுத்தல் Ser2 இல் நேரடி NRF40 பாஸ்போரிலேஷனை விளைவிக்கிறது, NRF2 கருவுக்கு இடமாற்றம் மற்றும் இலக்கு மரபணுக்களை செயல்படுத்துகிறது. IRE1 செயல்படுத்தல் TRAF2 இன் ஆட்சேர்ப்பைத் தூண்டுகிறது, அதைத் தொடர்ந்து ASK1 மற்றும் JNK பாஸ்போரிலேஷன் மற்றும் செயல்படுத்தல். JNK ஆனது பாஸ்போரிலேட் மற்றும் NRF2 ஐ செயல்படுத்துவதாக அறிவிக்கப்பட்டதால், IRE1 செயல்படுத்தல் NRF2 செயல்பாட்டை அதிகரிக்க வழிவகுக்கும் என்று நினைப்பது நியாயமானது.
UPR இன் தூண்டல் பற்றிய பல ஆய்வுகள் புரோட்டீன் கிளைகோசைலேஷன் துனிகாமைசின் தடுப்பானுடன் நடத்தப்பட்டுள்ளன. துனிகாமைசின்-தூண்டப்பட்ட அப்போப்டொடிக் செல் இறப்பைத் தடுப்பதற்கு NRF2 இன்றியமையாததாகத் தோன்றுகிறது [31] மேலும் இந்த நிலைமைகளின் கீழ் அதன் செயல்பாடு KEAP1 [51] இன் தன்னியக்க சிதைவால் இயக்கப்படுகிறது. அதன்படி, ?TC-2 செல்களில் NRF6 வெளிப்பாட்டின் shRNA-மத்தியஸ்த அமைதிப்படுத்தல், ஒரு முரைன் இன்சுலினோமா ?-செல் லைன், துனிகாமைசின் தூண்டப்பட்ட சைட்டோடாக்சிசிட்டியை கணிசமாக அதிகரித்தது மற்றும் அபோப்டோடிக் சார்பு ER ஸ்ட்ரெஸ் மார்க்கர் CHOP10 இன் வெளிப்பாட்டின் அதிகரிப்புக்கு வழிவகுத்தது. மறுபுறம், 2-டிதியோல்-1,2-தியோன் (D3T) மூலம் NRF3 செயல்படுத்தல் துனிகாமைசின் சைட்டோடாக்சிசிட்டியைக் குறைத்தது மற்றும் CHOP10 மற்றும் PERK [52] ஆகியவற்றின் வெளிப்பாட்டைக் குறைக்கிறது. சுவாரஸ்யமாக, டியூனிகாமைசினின் முறையான பயன்பாட்டிற்கு சமர்ப்பிக்கப்பட்ட ஆல்ஃபாக்டரி நியூரான்கள் CHOP, BIP, XBP2 [1] போன்ற மற்ற UPR-உறுப்பினர்களுடன் இணையாக NRF53 ஐ அதிகரித்தன. இந்த முடிவுகள் vivo ஆய்வுகளில் நீட்டிக்கப்பட்டுள்ளன, ஏனெனில் எலிகளில் tunicamycin இன் பக்கவாட்டு வென்ட்ரிகுலர் உட்செலுத்துதல் ஹிப்போகாம்பஸில் PERK மற்றும் NRF2 ஆகியவற்றின் வெளிப்பாட்டைத் தூண்டியது, குறிப்பிடத்தக்க அறிவாற்றல் குறைபாடுகள், அதிகரித்த TAU பாஸ்போரிலேஷன் மற்றும் A?42 வைப்புக்கள் [54].
NRF2 அப்-ஈஆர் உடலியலைப் பராமரிப்பதற்கான முக்கிய மரபணுக்களை ஒழுங்குபடுத்துகிறது
டைசல்பைட் வேதியியலைப் பராமரிக்க, ER லுமினுக்கு சைட்டோசோலில் இருந்து ஏராளமான GSH சப்ளை தேவைப்படுகிறது. NRF2 மூளையில் உள்ள ஜிஎஸ்ஹெச் வளர்சிதை மாற்றத்தின் முக்கியமான நொதிகளான சிஸ்டைன்/குளுட்டமேட் டிரான்ஸ்போர்ட், ?-குளூட்டமேட் சிஸ்டைன் சின்தேடேஸ் (?-ஜிஎஸ்), குளுட்டமேட்-சிஸ்டைன் லிகேஸ் கேடலிடிக் மற்றும் மாடுலேட்டர் சப்யூனிட்கள் (ஜிசிஎல்சி மற்றும் ஜிசிஎல்டியோன்ஜி) மற்றும் gaselutathioneG ரீடக்ட் போன்றவை. குளுதாதயோன் பெராக்சிடேஸ் (GPX) ([55] இல் மதிப்பாய்வு செய்யப்பட்டது). ER இல் GSH ஐ பராமரிப்பதில் NRF2 இன் பொருத்தம், NRF2 இன் மருந்தியல் அல்லது மரபணு செயலாக்கம் GCLC/GCLM வழியாக GSH தொகுப்பை அதிகரிப்பதைக் கண்டுபிடிப்பதன் மூலம் ஆதரிக்கப்படுகிறது, அதே நேரத்தில் NRF2-நாக் டவுன் மூலம் இந்த நொதிகளின் வெளிப்பாட்டைத் தடுப்பது சேதமடைந்த திரட்சியை ஏற்படுத்தியது. ER க்குள் உள்ள புரதங்கள் UPR செயல்படுத்தலுக்கு வழிவகுக்கும் [56].
C. elegans இல் Ire1, Xbp1 மற்றும் Atf1 உட்பட SKN-6 ஆல் கட்டுப்படுத்தப்படும் UPR இலக்கு மரபணுக்களின் பல கூறுகள். பாலூட்டிகளில் பல பெராக்சிடேஸ் (PRX) மற்றும் குளுதாதயோன் பெராக்சிடேஸ் (GPX) மரபணுக்களின் வெளிப்பாட்டை NRF2 மேல்-ஒழுங்குபடுத்தினாலும் ([57] இல் மதிப்பாய்வு செய்யப்பட்டது), GPX8 மட்டுமே ஒரு நேர்மையான ER-உள்ளூர்மயமாக்கப்பட்ட என்சைம் ஆகும், இது KDEL மீட்டெடுப்பு சிக்னலைக் கொண்டுள்ளது [58]. GPX8 இன் இழப்பு, UPR செயல்படுத்தலை ஏற்படுத்துகிறது, ERO1?-உருவாக்கப்பட்ட ஹைட்ரஜன் பெராக்சைடு சைட்டோசோலுக்கு கசிவு மற்றும் செல் இறப்பை ஏற்படுத்துகிறது. ஹைட்ரஜன் பெராக்சைடு ERO1 இலிருந்து பெறப்பட்டதா? GPX8 மற்றும் PRX4 [59] ஆகியவற்றின் ஒருங்கிணைந்த செயல்பாட்டின் காரணமாக செயல்பாடு ER இலிருந்து சைட்டோசோலுக்கு பரவ முடியாது. இது சம்பந்தமாக, வன வகை மற்றும் NRF2-பூஜ்ய எலி திசுக்களில் இருந்து RNA ஐப் பயன்படுத்தி ஆக்ஸிஜனேற்ற பாதுகாப்பு பாதை-மரபணு வெளிப்பாடு வரிசையின் பகுப்பாய்வு, NRF8 இல்லாத நிலையில் GPX2 இன் வெளிப்பாடு குறைவாக கட்டுப்படுத்தப்பட்டது என்பதை வெளிப்படுத்தியது [60]. இதற்கு இணங்க, மைலோப்ரோலிஃபெரேடிவ் நியோபிளாம்கள், பாலிசித்தீமியா அல்லது மைலோஃபைப்ரோஸிஸ், நோய்களும் ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் குறைந்த தர நாட்பட்ட அழற்சி ஆகியவற்றால் பாதிக்கப்பட்ட நோயாளிகளின் மாதிரிகளிலிருந்து டிரான்ஸ்கிரிப்டோம் பகுப்பாய்வு, கட்டுப்பாட்டுப் பாடங்களுடன் ஒப்பிடும்போது NRF2 மற்றும் GPX8 இரண்டின் வெளிப்பாடு அளவுகள் குறைவாக இருப்பதைக் காட்டுகிறது [61]. மனித மூளைப் பாதுகாப்பில் குறிப்பாக GPX8 ஐ உள்ளடக்கிய ஆய்வுகள் இன்னும் இல்லை, ஆனால் எலிகளில் ஒரு டிரான்ஸ்கிரிப்டோம் பகுப்பாய்வு பார்கின்சோனியன் நச்சு MPTP க்கு பதில் GPX8 அதிகரிப்பைக் குறிக்கிறது [62].
நியூரோடிஜெனரேட்டிவ் நோய்களில் UPR ஒழுங்குபடுத்தலில் NRF2 இன் தாக்கம்
PDI என்சைம்களின் செயலிழப்பு மற்றும் UPR இன் நாள்பட்ட செயல்படுத்தல் ஆகியவை நியூரோடிஜெனரேஷனைத் தொடங்கலாம் அல்லது துரிதப்படுத்தலாம். நோயால் பாதிக்கப்பட்ட நியூரான்கள், நியூரோடிஜெனரேட்டிவ் நோயின் விலங்கு மாதிரிகள் மற்றும் பிரேத பரிசோதனை செய்யப்பட்ட மனித திசுக்கள் இந்த கோளாறுகளில் பெரும்பாலானவற்றில் பல UPR-குறிப்பான்களின் மேல்-ஒழுங்குமுறைக்கு சான்று. நியூரோடிஜெனரேட்டிவ் நோய்களில் PDI/UPR பாதையின் மாற்றம் [63] இல் நன்றாக மதிப்பாய்வு செய்யப்பட்டுள்ளது, ஆனால் மூளையின் பிரேத பரிசோதனை மாதிரிகளில் இருந்து பின்வரும் சிறப்பம்சங்களைக் கருத்தில் கொள்ள வேண்டும். சிக்கலைத் தாங்கும் நியூரான்கள் மற்றும் AD மற்றும் PD நோயாளிகளின் லூயி உடல்களில் முறையே [64], [65] PDI அளவுகள் அதிகரிக்கப்படுகின்றன. PDI மற்றும் ERP57 ஆகியவை CSF இல் ALS நோயாளிகளிடமிருந்தும், CJD பாடங்களில் இருந்து மூளையிலும் கட்டுப்படுத்தப்படுகின்றன [66], [67], [68]. AD, PD அல்லது ALS [1], [6], [69], [70] நோயாளிகளின் மாதிரிகளில் BIP, PERK, IRE71 மற்றும் ATF67 ஆகியவை உயர்த்தப்பட்டுள்ளன. BIP, CHOP மற்றும் XBP1 ஆகியவை HD [72], [73] இலிருந்து பிரேத பரிசோதனை மூளை மாதிரிகளில் உயர்த்தப்படுகின்றன. மேலும், CJD நோயாளிகளிடமிருந்து கார்டெக்ஸ் திசுக்களில் ERP57, GRP94 மற்றும் BIP ஆகியவற்றின் மேல்-கட்டுப்பாடு கண்டறியப்பட்டது [74]. ஒட்டுமொத்தமாக, மூளையின் பாரன்கிமாவில் தவறாக மடிக்கப்பட்ட புரதங்களின் குவிப்பு UPR இன் தீங்கு விளைவிக்கும் மற்றும் நாள்பட்ட செயல்பாட்டிற்கு வழிவகுக்கிறது என்பதை இந்த ஆதாரம் வெளிப்படுத்துகிறது. சுவாரஸ்யமாக, கி.பி தொடக்கத்தில் PERK ஆல் NRF2 செயல்படுத்துவதை இணைக்கும் சமீபத்திய ஆய்வு உள்ளது. இந்த ஆய்வில், பல்வேறு நோய் நிலைகளில் மனித புற இரத்த அணுக்கள் மற்றும் ஒரு AD டிரான்ஸ்ஜெனிக் மவுஸ் மாதிரியைப் பயன்படுத்தி AD நோய்க்கிருமிகளின் ஆரம்ப நிகழ்வுகளை NRF2 மற்றும் UPR ஆகியவற்றில் ஆக்ஸிஜனேற்ற அழுத்த மத்தியஸ்த மாற்றங்கள் உருவாக்க முடியுமா என்பதை ஆசிரியர்கள் ஆய்வு செய்தனர். லேசான அறிவாற்றல் குறைபாடு உள்ள நபர்களிடமிருந்து தனிமைப்படுத்தப்பட்ட மனித புற இரத்த மோனோநியூக்ளியர் செல்களில் அதிகரித்த ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் அதிகரித்த pSer40-NRF2 ஆகியவை காணப்பட்டன. மேலும், லேசான அறிவாற்றல் குறைபாடு மற்றும் லேசான AD [75] உள்ள நபர்களிடமிருந்து இந்த செல்களில் பலவீனமான ER கால்சியம் ஹோமியோஸ்டாஸிஸ் மற்றும் மேல்-ஒழுங்குபடுத்தப்பட்ட ER- அழுத்த குறிப்பான்கள் இருப்பதாக அவர்கள் தெரிவித்தனர்.
NRF2 மற்றும் Ubiquitin Proteasome'System (UPS) ஆகியவற்றின் பரஸ்பர ஒழுங்குமுறை
யுபிஎஸ் NRF2 புரத நிலைகளை மாற்றியமைக்கிறது
UPS ஆனது சேதமடைந்த அல்லது தவறாக மடிக்கப்பட்ட புரதங்களின் சிதைவில் பங்கேற்கிறது மற்றும் சைட்டோசோல் மற்றும் நியூக்ளியஸில் உள்ள முக்கிய ஒழுங்குமுறை மூலக்கூறுகளின் அளவைக் கட்டுப்படுத்துகிறது. இந்த அமைப்பின் மைய மையமானது ஒரு பெரிய மல்டிசபுனிட் என்சைம் ஆகும், இது 20S எனப்படும் புரோட்டியோலிடிக் செயலில் உள்ள வளாகத்தைக் கொண்டுள்ளது. 20S கோர் புரோட்டீசோம் விரிந்த புரதங்களை சிதைக்கிறது, ஆனால் வெவ்வேறு ஒழுங்குமுறை புரத வளாகங்களுடன் பிணைப்பது அதன் அடி மூலக்கூறு விவரக்குறிப்பு மற்றும் செயல்பாட்டை மாற்றுகிறது. உதாரணமாக, 19S மையத்தில் ஒன்று அல்லது இரண்டு 20S ஒழுங்குமுறை துணைப்பிரிவுகளைச் சேர்ப்பது 26S புரோட்டீசோமை உருவாக்குகிறது மற்றும் நேட்டிவ் மடிந்த புரதங்களை நோக்கி அதன் தனித்தன்மையை மாற்றுகிறது [76], [77]. புரோட்டீசோமால் சிதைவுக்கு எபிக்விடின் கோவலன்ட் பிணைப்பு தேவைப்படுகிறது. எபிக்விடின் இணைத்தல் மூன்று-படி அடுக்கு பொறிமுறையின் வழியாக தொடர்கிறது. முதலாவதாக, எபிக்விடின்-செயல்படுத்தும் என்சைம் E1, ஏடிபி தேவைப்படும் எதிர்வினையில் எபிக்விடினை செயல்படுத்துகிறது. பின்னர், ஒரு E2 என்சைம் (ubiquitin-carrier protein அல்லது ubiquitin-conjugating enzyme) செயல்படுத்தப்பட்ட ubiquitin ஐ E1 இலிருந்து E3 என பெயரிடப்பட்ட ubiquitin-protein ligase குடும்பத்தின் உறுப்பினருடன் இணைக்கப்பட்ட அடி மூலக்கூறுக்கு மாற்றுகிறது. எங்கும் நிறைந்த-புரதத்தின் சரியான விதி எபிக்விடின் சங்கிலியின் தன்மையைப் பொறுத்தது என்றாலும், இந்த செயல்முறை பொதுவாக 26S புரோட்டீசோம் [78] மூலம் சிதைவை ஏற்படுத்துகிறது.
E3-ligase KEAP1 என்பது NRF2 இன் சிறந்த அறியப்பட்ட தடுப்பானாகும். KEAP1 ஒழுங்குமுறையின் பொறிமுறையானது NRF2 அளவுகள் ஆக்ஸிஜனேற்ற ஏற்ற இறக்கங்களுக்கு எவ்வாறு சரிசெய்கிறது என்பதை நேர்த்தியாக விளக்குகிறது. அடிப்படை நிலைமைகளின் கீழ், புதிதாக ஒருங்கிணைக்கப்பட்ட NRF2 ஹோமோடைமர் KEAP1 ஆல் கைப்பற்றப்படுகிறது, இது ஒரு NRF2 மூலக்கூறை இரண்டு அமினோ அமில வரிசைகளில் குறைந்த (அஸ்பார்டேட், லுசின், கிளைசின்; DLG) மற்றும் உயர் (குளுட்டமேட், த்ரோயோனைன், கிளைசின், ETGEmate) உடன் பிணைக்கிறது. KEAP1 உடனான தொடர்பு NRF2 ஐ CULIN3/RBX1 புரத வளாகத்திற்கு வழங்க உதவுகிறது, இதன் விளைவாக அதன் எங்கும் பரவுதல் மற்றும் அதைத் தொடர்ந்து புரோட்டீசோமால் சிதைவு ஏற்படுகிறது. இருப்பினும், KEAP1 இன் ரெடாக்ஸ் மாற்றமானது CULIN2/RBX3 ஆல் குறிப்பிடப்படும் UPS க்கு NRF1 வழங்குவதைத் தடுக்கிறது. இதன் விளைவாக, புதிதாக ஒருங்கிணைக்கப்பட்ட NRF2 KEAP1-சார்ந்த சிதைவிலிருந்து தப்பித்து, கருவில் குவிந்து ARE-கொண்ட மரபணுக்களை [79], [80], [81], [82] செயல்படுத்துகிறது.
E3-லிகேஸ் அடாப்டர் ?-TrCP ஆனது GSK-2? மூலம் NRF3 இன் பாஸ்போரிலேஷன் தொடர்பான சமிக்ஞை நிகழ்வுகளில் பங்கேற்கும் ஹோமோடைமர் ஆகும். இந்த கைனேஸ் NRF2 (aspartate, serine, glycine, isoleucine serine; DSGIS) இன் குறிப்பிட்ட செரின் எச்சங்களை பாஸ்போரிலேட் செய்து ஒரு சிதைவு டொமைனை உருவாக்குகிறது, அது ?-TrCP ஆல் அங்கீகரிக்கப்பட்டு CULLIN1/RBX1 காம்ப்ளக்ஸ் மூலம் புரோட்டீசோம் சிதைவுக்கு குறியிடப்படுகிறது. GSK-3 மூலம் பாஸ்போரிலேட்டட் செய்யப்பட்ட குறிப்பிட்ட அமினோ அமிலங்களின் அடையாளம்? Neh6 டொமைன், 2D-ஜெல் எலக்ட்ரோபோரேசிஸ் [15], [26] மற்றும் மாஸ் ஸ்பெக்ட்ரோஸ்கோபி [83] ஆகியவற்றின் தளம்-இயக்கப்பட்ட பிறழ்வுகளின் கலவையால் இந்த டிக்ரோன் நடத்தப்பட்டது. இதன் விளைவாக, GSK-3 இன் தடுப்பு? GSK-3 ஐசோஃபார்ம்களுக்கு எதிராக மிகவும் தேர்ந்தெடுக்கப்பட்ட மருந்துகள் அல்லது siRNAகள் மூலம் NRF2 புரத அளவுகள் அதிகரித்தன. ?-TrCP ஐசோஃபார்ம்கள் 1 மற்றும் 2 க்கு எதிரான siRNA களில் இதே போன்ற முடிவுகள் காணப்பட்டன. GSK-2 ஐத் தொடர்ந்து NRF3 இன் நிலைப்படுத்தல்? KEAP1-குறைபாடுள்ள மவுஸ் எம்ப்ரியோ ஃபைப்ரோபிளாஸ்ட்களில் தடுப்பு ஏற்பட்டது மற்றும் ஒரு எக்டோபிகல் வெளிப்படுத்தப்பட்ட NRF2 நீக்குதல் விகாரியில் KEAP1 உடன் உயர்-தொடர்பு பிணைப்புக்கான முக்கியமான ETGE எச்சங்கள் இல்லாதது, மேலும் KEAP1-சுயாதீனமான ஒழுங்குமுறையை நிரூபிக்கிறது.
நியூரோடிஜெனரேட்டிவ் நோய்களின் பின்னணியில், யுபிஎஸ் மூலம் NRF2 இன் பண்பேற்றத்தை இரண்டு வெவ்வேறு வழிகளில் நாம் கற்பனை செய்யலாம். ஒருபுறம், KEAP1 அமைப்பு தவறாக மடிக்கப்பட்ட புரதக் குவிப்பிலிருந்து பெறப்பட்ட ரெடாக்ஸ் ஏற்றத்தாழ்வை உணரும், அதே சமயம் GSK-3/?-TrCP அச்சு புரோட்டியோஸ்டாசிஸ் இழப்பால் மாற்றப்பட்ட சமிக்ஞை கடத்துதலில் ஒரு செயலில் பங்கேற்பாளராக செயல்படும் (படம். 2).
படம் 2 UPS ஆனது NRF2 அளவுகளை இறுக்கமாக கட்டுப்படுத்துகிறது. ஹோமியோஸ்ட்டிக் நிலைமைகளின் கீழ், குறைந்த NRF2 அளவுகள் E3 லிகேஸ் அடாப்டர்கள் KEAP1 மற்றும் ?-TrCP ஆகியவற்றின் செயல்பாட்டால் பராமரிக்கப்படுகின்றன. இடதுபுறத்தில், NRF2 குறைந்த (DLG) மற்றும் உயர் (ETGE) தொடர்பு மையக்கருத்துகள் மூலம் KEAP1 ஹோமோடைமரின் கெல்ச் டொமைன்களுடன் பிணைக்கிறது. அதன் BTB டொமைன் மூலம், KEAP1 ஒரே நேரத்தில் CULIN3/RBX1 வளாகத்துடன் பிணைக்கிறது, NRF2 எங்கும் பரவுதல் மற்றும் 26 S புரோட்டீசோம் மூலம் சிதைவைச் செயல்படுத்துகிறது. மேலும், GSK-3? ஒரு சிதைவு டொமைனை (DpSGIpSL) உருவாக்க NRF335 இன் பாஸ்போரிலேட்டுகள் Ser338 மற்றும் Ser2 எச்சங்களை உருவாக்குகிறது, இது ubiquitin ligase adapter ?-TrCP ஆல் அங்கீகரிக்கப்பட்டு CULIN3/RBX1 வளாகத்தால் புரோட்டீசோம் சிதைவுக்கு குறியிடப்பட்டது. சரி, வினைத்திறன் ஆக்சிஜன் இனங்கள் அல்லது எலெக்ட்ரோஃபைல்களின் வெளிப்பாட்டின் போது KEAP1 இல் முக்கியமான Cys எச்சங்கள் மாற்றியமைக்கப்படுகின்றன, KEAP1 ஆனது NRF2 அல்லது CULIN3/RBX1 உடன் திறம்பட தொடர்பு கொள்ள முடியாது, பின்னர் இந்த டிரான்ஸ்கிரிப்ஷன் காரணி ARE-மரபணுக்களை நோக்கி அதன் அரை-வாழ்க்கை மற்றும் டிரான்ஸ்கிரிப்ஷனல் செயல்பாட்டை அதிகரிக்கிறது. GSK-3 ஐத் தடுக்கும் சிக்னலிங் பாதைகள், Ser9 இல் உள்ள AKT பாஸ்போரிலேஷன், இலக்கு மரபணுக்களின் புரோட்டீசோம், குவிப்பு மற்றும் தூண்டல் ஆகியவற்றால் NRF2 சிதைவை ஏற்படுத்துகிறது.
NRF2 புரோட்டீசோம் துணைக்குழுக்களின் டிரான்ஸ்கிரிப்ஷனல் கட்டுப்பாடு மூலம் யுபிஎஸ் செயல்பாட்டை அதிகரிக்கிறது
NRF2 பல புரோட்டீசோம் துணைக்குழுக்களின் வெளிப்பாட்டைக் கட்டுப்படுத்துகிறது, இதனால் செல்களை நச்சு புரதங்களின் திரட்சியிலிருந்து பாதுகாக்கிறது. இருபது புரோட்டீசோம்- மற்றும் எங்கும் பரவுதல் தொடர்பான மரபணுக்கள் NRF2 ஆல் கட்டுப்படுத்தப்படுவதாகத் தோன்றுகிறது, இது கல்லீரல் ஆர்என்ஏவிலிருந்து ஒரு பரந்த மைக்ரோஅரே பகுப்பாய்வின் படி NRF2 தூண்டி D3T [84] உடன் அமைக்கப்பட்டது. ஒரு பின்பக்க ஆய்வில், அதே ஆசிரியர்கள் 26S புரோட்டீசோமின் பெரும்பாலான துணைப்பிரிவுகளின் வெளிப்பாடு D3T உடன் சிகிச்சையளிக்கப்பட்ட எலிகளிலிருந்து கல்லீரலில் மூன்று மடங்கு வரை மேம்படுத்தப்பட்டதாகக் காட்டியுள்ளனர். சப்யூனிட் புரத அளவுகள் மற்றும் புரோட்டீசோம் செயல்பாடு ஒருங்கிணைக்கப்பட்டது. இருப்பினும், டிரான்ஸ்கிரிப்ஷன் காரணி NRF2 சீர்குலைந்த எலிகளில் எந்த தூண்டுதலும் காணப்படவில்லை. PSMB5 (20S) புரோட்டீசோம் சப்யூனிட்டின் ஊக்குவிப்பு செயல்பாடு NRF2 அதிகப்படியான அழுத்தம் அல்லது மவுஸ் எம்ப்ரியோனிக் ஃபைப்ரோபிளாஸ்ட்களில் ஆக்டிவேட்டர்களுடன் சிகிச்சையளிப்பதன் மூலம் அதிகரித்தது, மேலும் PSMB5 [85] இன் அருகாமை ஊக்குவிப்பாளரில் AREகள் அடையாளம் காணப்பட்டன. NRF2 இன் மருந்தியல் செயலாக்கமானது, செயல்பாட்டு NRF3 [6] கொண்ட முதிர்ச்சியற்ற மனித ஃபைப்ரோபிளாஸ்ட்களில் மட்டுமே பிரதிநிதித்துவ புரோட்டீசோம் துணைக்குழுக்களின் (PSMA1, PSMA5, PSMB2 மற்றும் PSMB86) வெளிப்பாடு நிலைகளை உயர்த்தியது. ஆக்ஸிஜனேற்ற அழுத்தத்திற்குத் தழுவலின் போது NRF2 செயல்படுத்தல் PSMB1 (20S) மற்றும் PA28 இன் உயர் வெளிப்பாட்டை விளைவிக்கிறது? துணை அலகுகள் (அல்லது S11, புரோட்டீசோம் ரெகுலேட்டர்) [87]. மேலும், மனித கரு ஸ்டெம் செல்களின் முடிவுகள், புரோட்டீசோம் முதிர்வு புரதத்தின் (POMP) வெளிப்பாட்டை NRF2 கட்டுப்படுத்துகிறது, இது ஒரு புரோட்டீசோம் சேப்பரோன், இது சுய-புதுப்பிக்கும் மனித கரு ஸ்டெம் செல்களின் பெருக்கத்தை மாற்றியமைக்கிறது, மூன்று கிருமி அடுக்கு வேறுபாடு மற்றும் செல்லுலார் மறுசீரமைப்பு [ 88]. இந்த ஆய்வுகள் அனைத்தும் சேர்ந்து, NRF2 UPS இன் முக்கிய கூறுகளின் வெளிப்பாட்டை அதிக-ஒழுங்குபடுத்துகிறது, எனவே நச்சுத்தன்மையுள்ள புரதங்களை அகற்றுவதில் தீவிரமாக பங்களிக்கிறது.
நியூரோடிஜெனரேட்டிவ் நோய்களில் NRF2-UPS அச்சு
நரம்பியக்கடத்தல் நோய்களில் UPS இன் பங்கு தீவிர விவாதத்தின் ஒரு துறையாகும். ஆரம்ப ஆய்வுகள் பல நரம்பியக்கடத்தல் நோய்களால் பாதிக்கப்பட்ட நோயாளிகளின் மனித நரபலிகளில் புரோட்டீசோம் செயல்பாடு குறைந்துவிட்டதாக அறிவித்தது. இருப்பினும், விட்ரோ மற்றும் விவோ அணுகுமுறைகளைப் பயன்படுத்தும் பிற ஆய்வுகள் மாறாமல் அல்லது அதிகரித்த புரோட்டீசோம் செயல்பாட்டைக் கண்டறிந்தன ([89] இல் மதிப்பாய்வு செய்யப்பட்டது). இந்த முரண்பாட்டிற்கான ஒரு சாத்தியமான விளக்கம் என்னவென்றால், நோயின் வளர்ச்சியின் போது UPS கூறுகளின் அளவுகள் மாறலாம் மற்றும் NRF2-இலக்குகளுக்கு பரிந்துரைக்கப்பட்டதைப் போல வெவ்வேறு மூளைப் பகுதிகளில் மாறலாம்.
இந்த சர்ச்சை இருந்தபோதிலும், ARE-கொண்ட புரோட்டீசோம் மரபணுக்களின் மேல்-கட்டுப்பாடு மூளையில் உள்ள நச்சு புரதங்களின் அனுமதியை அதிகரிப்பதன் மூலம் UPS ஐ வலுப்படுத்தும் என்பதை கவனத்தில் கொள்ள வேண்டும். உண்மையில், நரம்பணு உயிரணுக்களில் ஆன்டிஆக்ஸிடன்ட் பதிலின் மாடுலேட்டரான NRF1 ஐ நீக்குவது, புரோட்டீசோம் செயல்பாடு மற்றும் நரம்பியக்கடத்தல் குறைபாடுகளுக்கு வழிவகுக்கிறது. குரோமாடின் இம்யூனோபிரெசிபிட்டேஷன் பரிசோதனைகள் மற்றும் டிரான்ஸ்கிரிப்ஷனல் பகுப்பாய்வு PSMB6 NRF1 ஆல் கட்டுப்படுத்தப்படுகிறது என்பதை நிரூபித்தது. கூடுதலாக, மரபணு வெளிப்பாடு விவரக்குறிப்பு நியூரான்களில் உள்ள புரோட்டீசோம் மரபணுக்களின் முக்கிய டிரான்ஸ்கிரிப்ஷனல் ரெகுலேட்டராக NRF1 ஐ அடையாளம் காண வழிவகுத்தது, NRF1 இல் உள்ள இடையூறுகள் நரம்பியக்கடத்தல் நோய்களின் நோய்க்கிருமி உருவாக்கத்திற்கு பங்களிக்கக்கூடும் என்று பரிந்துரைக்கிறது [90]. சுவாரஸ்யமாக, NRF1 மற்றும் TCF11 எனப்படும் அதன் நீண்ட ஐசோஃபார்ம், குறைக்கப்பட்ட புரோட்டியோலிடிக் செயல்பாட்டை ஈடுசெய்ய ஒரு பின்னூட்ட சுழற்சியில் புரோட்டீசோம் தடுப்பின் மீது ARE-கொண்ட புரோட்டீசோம் மரபணுக்களை கட்டுப்படுத்துவதாகக் காட்டப்பட்டது [91], [92].
NRF2 ஐப் பொறுத்தவரை, DJ-2-குறைபாடுள்ள எலிகளின் நடுமூளையில் NRF6, RPT19 (5 S) மற்றும் PSMB20 (1 S) அளவுகளைக் குறைப்பதில் ஒரு தொடர்பு உள்ளது [93]. மேலும், இயற்கையாக நிகழும் சல்ஃபோராபேன் (SFN) கலவையானது UPS இன் முக்கியமான மாடுலேட்டராக NRF2 இன் மிகவும் வலுவான படத்தை அளிக்கிறது. முரைன் நியூரோபிளாஸ்டோமா நியூரோ2ஏ செல்கள் கொண்ட விட்ரோ சோதனைகளில் புரோட்டீசோமின் வினையூக்கி துணைக்குழுக்களின் மேம்பட்ட வெளிப்பாட்டையும், SFN க்கு பதிலளிக்கும் வகையில் அதன் பெப்டிடேஸ் செயல்பாடுகளையும் நிரூபிக்கிறது. இந்த மருந்து ஹைட்ரஜன் பெராக்சைடு-மத்தியஸ்த சைட்டோடாக்ஸிசிட்டி மற்றும் புரோட்டீசோம் செயல்பாட்டைச் சார்ந்து புரத ஆக்சிஜனேற்றத்திலிருந்து செல்களைப் பாதுகாத்தது [94]. கூடுதலாக, லியு மற்றும் சக பணியாளர்கள் மூளையில் உள்ள SFN க்கு பதிலளிக்கும் வகையில் UPS செயல்பாட்டை கண்காணிக்க ஒரு நிருபர் சுட்டியைப் பயன்படுத்தினார்கள். UPS (GFPu) மூலம் அதன் விரைவான சீரழிவை ஊக்குவிக்கும் ஒரு கட்டமைப்பு சிதைவு சமிக்ஞையுடன் இணைந்த பச்சை ஒளிரும் புரதத்தை (GFP) இந்த எலிகள் எங்கும் வெளிப்படுத்துகின்றன. பெருமூளைப் புறணியில், 5 எஸ் புரோட்டீசோமின் சைமோட்ரிப்சின் போன்ற (PSMB2), காஸ்பேஸ் போன்ற (PSMB1) மற்றும் டிரிப்சின் போன்ற (PSMB20) செயல்பாடுகளில் இணையான அதிகரிப்புடன் GFPu இன் அளவை SFN குறைத்தது. கூடுதலாக, SFN உடன் ஹண்டிங்டன்-பெறப்பட்ட செல்கள் சிகிச்சையானது NRF2 செயல்படுத்தல் mHtt சிதைவை மேம்படுத்தியது மற்றும் mHtt சைட்டோடாக்சிசிட்டியைக் குறைத்தது [95]. SFN செயல்பாட்டின் முக்கிய வழிமுறையானது NRF2 [96] இன் தூண்டல் மூலமாகும். NRF2 இன் குறிப்பிட்ட பங்களிப்பானது மேலதிக ஆய்வுகளில் NRF2-பூஜ்ய அமைப்புகளைப் பயன்படுத்துவதன் மூலம் கவனிக்கப்பட வேண்டும்.
NRF2 மற்றும் மேக்ரோஆட்டோபாகி இடையே செயல்பாட்டு இணைப்பு
NRF2 புரத அளவுகள் அடாப்டர் புரோட்டீன் P62 மூலம் மாற்றியமைக்கப்படுகின்றன
ஆட்டோபேஜி என்பது லைசோசோம்களுக்குள் உள்ள சைட்டோசோலிக் கூறுகளின் சிதைவைக் குறிக்கிறது. இந்த செயல்முறை நீண்ட கால மற்றும் தவறாக மடிந்த புரதங்கள் மற்றும் சேதமடைந்த உறுப்புகளை அகற்ற பயன்படுகிறது. NRF2 மற்றும் தன்னியக்கத்திற்கு இடையேயான நேரடி இணைப்பு, அடாப்டர் புரதம் p62 உடன் முதலில் காணப்பட்டது, இது SQSTM1 [97], [98], [99], [100], [101] என்றும் அழைக்கப்படுகிறது. இந்த புரதம் எங்கும் பரவும் புரதங்களை புரோட்டீசோமால் மற்றும் லைசோசோமால் சிதைவு இயந்திரங்களுக்கு அனுப்புகிறது மற்றும் சேதமடைந்த புரதங்களை அவற்றின் சிதைவுக்கு முன் மொத்தமாக பிரிக்கிறது. P62 ஆனது எங்கும் பரவும் புரதங்களுடன் பிணைப்பதற்காக எபிக்விடின்-தொடர்புடைய (UBA) டொமைனையும், தன்னியக்க ஏற்பி LC3 மூலம் தன்னியக்க சவ்வுடன் ஒருங்கிணைக்க LC3-இன்டராக்டிங் பிராந்தியத்தையும் (LIR) வழங்குகிறது.
NRF62 மற்றும் அதன் இலக்கு மரபணுக்களின் p2-மத்தியஸ்த தூண்டல் முதன்முதலில் 2007 இல் தெரிவிக்கப்பட்டது [102], KEAP1 [103], [98], [99], [100] உடனான அதன் தொடர்பு கண்டுபிடிக்கும் வரை மூலக்கூறு வழிமுறை முழுமையாகப் புரிந்து கொள்ளப்படவில்லை. ], [101]. கோமாட்சு மற்றும் சக பணியாளர்கள் P1 இல் KEAP62 ஊடாடும் பகுதியை (KIR) அடையாளம் கண்டுள்ளனர், அது KEAP1 ஐ NRF2 போன்ற அதே அடிப்படை மேற்பரப்புப் பாக்கெட்டில் பிணைத்தது மற்றும் NRF2 இல் ETGE மையக்கருத்தைப் போன்ற பிணைப்புத் தொடர்பைக் கொண்டுள்ளது, இது p62 மற்றும் NRF2 இடையே போட்டியைக் குறிக்கிறது. P351 (62-DPSTGE-349) இல் உள்ள KIR மையக்கருத்தில் Ser354 இன் பாஸ்போரிலேஷன் KEAP1 க்கான அதன் தொடர்பை அதிகரிப்பதாகக் காட்டப்பட்டது, NRF2 பிணைப்புடன் போட்டியிடுகிறது மற்றும் அதன் இலக்கு மரபணுக்களின் குவிப்பு மற்றும் டிரான்ஸ்கிரிப்ஷனல் செயல்படுத்தலை அனுமதிக்கிறது [98], [99]. உண்மையில், p62 அதிகப்படியான வெளிப்பாடு NRF2 எங்கும் பரவுவதைக் குறைக்க வழிவகுத்தது மற்றும் அதன் விளைவாக உறுதிப்படுத்தல் மற்றும் அதன் இலக்கு மரபணுக்களின் தூண்டல் [104]. சில கைனேஸ்கள் p62 பாஸ்போரிலேஷனில் பங்கேற்க பரிந்துரைக்கப்பட்டுள்ளன. mTOR இன்ஹிபிட்டர் ராபமைசினுடனான சிகிச்சையானது p1 இன் பாஸ்போரிலேஷனை அடக்கியது மற்றும் ஆர்சனைட் சிகிச்சையின் போது KEAP1 இன் கீழ்-ஒழுங்குமுறை ஆகியவற்றை ரபமைசின் காம்ப்ளக்ஸ் 62 (mTORC1) இன் பாலூட்டிகளின் இலக்கு உட்படுத்தலாம். சமீபத்தில், TGF-?-செயல்படுத்தப்பட்ட கைனேஸ் 1 (TAK1) ஆனது p62 ஐ பாஸ்போரிலேட் செய்ய முடியும், இது KEAP1 சிதைவு மற்றும் NRF2-அதிக-ஒழுங்குமுறையை மேம்படுத்துகிறது. NRF1 புரத அளவுகள் குறைவதற்கு இணையாக வெவ்வேறு சுட்டி திசுக்களில் வெளிப்புற ஆக்சிஜனேற்றம் இல்லாத நிலையில் TAK2-குறைபாடு ROS ஐ ஒழுங்குபடுத்துவதால், நிலையான நிலையின் கீழ் செல்லுலார் ரெடாக்ஸ்டாசிஸை ஒழுங்குபடுத்துவதற்கான ஒரு வழி இது என்று இந்த ஆய்வின் ஆசிரியர்கள் பரிந்துரைக்கின்றனர் [105] ].
UBA டொமைன் இல்லாத ஒரு p62 கட்டமைப்பானது இன்னும் KEAP1 ஐ பிணைக்கும் திறன் கொண்டது, இது தொடர்பு எங்கும் பரவிய KEAP1 [101] ஐச் சார்ந்து இல்லை என்பதைக் குறிக்கிறது. இருப்பினும், டிரோசோபிலா மெலனோகாஸ்டரில் உள்ள p62 ஹோமோலாக், Ref(2) என பெயரிடப்பட்டது, KIR மையக்கருத்தைக் கொண்டிருக்கவில்லை மற்றும் DmKEAP1 உடன் நேரடியாக தொடர்பு கொள்ளாது, இருப்பினும் UBA டொமைன் மூலம் எங்கும் பரவிய DmKEAP1 உடன் பிணைக்க முடியும். மேலும், DmKEAP1 நேரடியாக Atg8 உடன் தொடர்பு கொள்ள முடியும் (பாலூட்டி LC3க்கு ஹோமோலாக்). KEAP1 குறைபாடு Atg8 மற்றும் தன்னியக்க தூண்டல் NRF2 ஆர்த்தோலாக் CncC மற்றும் TFEB/MITF [106] இல் சார்ந்துள்ளது. NRF2 மற்றும் தன்னியக்கத்திற்கு இடையிலான உறவு பாதுகாக்கப்பட்டதாகத் தெரிகிறது, இது அதன் செயல்பாட்டு பொருத்தத்தை எடுத்துக்காட்டுகிறது.
P2 ஆல் NRF62 இன் தூண்டல் ஆனது KEAP1 ஐ பிணைப்பதற்கான போட்டி மற்றும் லைசோசோமில் KEAP1 இன் சிதைவு ஆகிய இரண்டின் விளைவாகும். siRNA உடன் p62 ஐ அமைதிப்படுத்துவது NRF1 மற்றும் அதன் இலக்கு மரபணுக்கள் [2] குறைவதற்கு இணையாக KEAP101 அரை-வாழ்க்கை இரட்டிப்பாக்கியது. உடன்பாட்டில், காட்டு வகை எலிகளுடன் ஒப்பிடும்போது p62 வெளிப்பாட்டின் நீக்கம் KEAP1 இன் அளவு அதிகரித்தது. மிகவும் பொருத்தமானது, KEAP1 அளவுகளின் அதிகரிப்பு புரோட்டீசோம் தடுப்பான்களால் பாதிக்கப்படவில்லை, ஆனால் பட்டினியைத் தூண்டும் தன்னியக்கத்தின் கீழ் குறைக்கப்பட்டது [107]. உண்மையில், KEAP1 ஆனது p62 மற்றும் LC3 [99], [100], [103] ஆகியவற்றால் அலங்கரிக்கப்பட்ட தன்னியக்க வெசிகல்களில் உள்ள பாலூட்டிகளின் உயிரணுக்களில் உள்ளது. இந்தத் தரவுகள் அனைத்தும் KEAP1 என்பது மேக்ரோஆட்டோபாகி இயந்திரத்தின் அடி மூலக்கூறு என்று கூறுகின்றன, ஆனால் சில சர்ச்சைக்குரிய முடிவுகள் இருப்பதால் இந்த சிக்கலை இன்னும் விரிவாக பகுப்பாய்வு செய்ய வேண்டும். மேக்ரோஆட்டோபாகி [1] இன் முக்கிய விளைபொருளான Atg7-null எலிகளில் KEAP107 புரத அளவுகள் அதிகரிக்கப்பட்டன, ஆனால் torin1, E64/pepstatin அல்லது bafilomycin உடன் மேக்ரோஆட்டோபாகியின் மருந்தியல் தடுப்பு KEAP1 [107], [100] ஐக் குவிக்கத் தவறியது. ஒட்டுமொத்தமாக, இந்த முடிவுகள் அதிகரித்த p62 அளவுகள் KEAP1 ஐ தன்னியக்க வெற்றிடங்களாகப் பிரிப்பதாகக் கூறுகின்றன, மேலும் இது KEAP1 தன்னியக்கச் சிதைவில் NRF2 செயல்படுத்தலை அனுமதிக்கிறது (படம் 3). இந்த சூழலில் சல்பினிக் அமில ரிடக்டேஸ்கள் SESTRINS முக்கிய பங்கு வகிக்கிறது என்று இரண்டு வெவ்வேறு ஆய்வுகள் தெரிவிக்கின்றன. SESTRIN 2 ஆனது p62, KEAP1 மற்றும் RBX1 உடன் தொடர்பு கொள்கிறது மற்றும் இலக்கு மரபணுக்களை KEAP62 மற்றும் NRF1 செயல்படுத்தும் p2-சார்ந்த சிதைவை எளிதாக்குகிறது [108]. மற்றொரு ஆய்வு, SESTRIN 2 ஆனது ULK1 மற்றும் p62 உடன் தொடர்பு கொண்டு, Ser62 இல் p403 இன் பாஸ்போரிலேஷனை ஊக்குவிக்கிறது, இது KEAP1 [109] உள்ளிட்ட சரக்கு புரதங்களின் சிதைவை எளிதாக்கியது.
படம் 3 NRF2 அளவுகள் அடாப்டர் புரதம் p62 மூலம் கட்டுப்படுத்தப்படுகிறது. mTORC351, TAK62 அல்லது பிற கைனேஸ்கள் மூலம் p349 (354-DPSTGE-1) இன் KIR மையக்கருத்தில் Ser 1 இன் பாஸ்போரிலேஷன் NRF1 இல் உள்ள ETGE மையக்கருத்தை ஒத்திருப்பதன் காரணமாக KEAP2 உடன் பிணைப்பதில் அதிக ஈடுபாட்டை ஏற்படுத்துகிறது. இதன் விளைவாக, பாஸ்போரிலேட்டட் p62 NRF2 ஐ இடமாற்றம் செய்து KEAP1 ஐ பிணைக்கிறது. p62 இல் உள்ள LIR மையக்கருமானது தன்னியக்க சவ்வில் LC3 உடன் தொடர்பு கொள்ள உதவுகிறது, இதனால் p62-KEAP1 வளாகம் இறுதியில் லைசோசோமில் சிதைக்கப்படுகிறது. இதன் விளைவாக, NRF2 குவிந்து, அணுக்கருவுக்கு இடமாற்றம் செய்து, p62 உட்பட ARE-கொண்ட மரபணுக்களின் படியெடுத்தலை அதிகரிக்கிறது. NRF2 செயல்பாட்டைத் தடுப்பதற்காக KEAP1 புதிதாக ஒருங்கிணைக்கப்பட வேண்டியிருப்பதால், இந்த ஒழுங்குமுறை பொறிமுறையானது நீடித்த NRF2 பதிலை வழங்குகிறது.
NRF2 மூலம் மேக்ரோஆட்டோபாகி மரபணுக்களின் பண்பேற்றம்
NRF2 மேக்ரோஆட்டோபாகிக்கான தொடர்புடைய மரபணுக்களின் வெளிப்பாட்டை ஒழுங்குபடுத்துகிறது, அது UPR மற்றும் UPS க்கும் செய்கிறது. எலக்ட்ரோஃபைல்ஸ், ஆர்ஓஎஸ் மற்றும் நைட்ரிக் ஆக்சைடு [62], [110], [111] ஆகியவற்றின் வெளிப்பாட்டின் போது p112 வெளிப்பாடு தூண்டப்பட்டதாகக் காட்டப்பட்ட ஆய்வுகளிலிருந்து முதல் ஆதாரம் கிடைத்தது. தூண்டல் செயல்முறை சில ஆண்டுகளுக்குப் பிறகு p62 அதன் மரபணு ஊக்குவிப்பாளரில் ஒரு செயல்பாட்டு ARE ஐக் கொண்டுள்ளது என்பதைக் கண்டறிந்தது [99]. சமீபத்திய ஆய்வில், உயிர் தகவலியல் பகுப்பாய்வு மற்றும் சிஐபி மதிப்பீடுகளைத் தொடர்ந்து பல செயல்பாட்டு AREகள் கண்டறியப்பட்டு சரிபார்க்கப்பட்டன. மேலும், Nrf2-நாக் அவுட் எலிகளின் மவுஸ் எம்ப்ரியோனிக் ஃபைப்ரோபிளாஸ்ட்கள் மற்றும் கார்டிகல் நியூரான்கள் குறைக்கப்பட்ட p62 வெளிப்பாட்டை வெளிப்படுத்தின, இது NRF2-எக்ஸ்பிரஸிங் லென்டிவைரஸ் மூலம் மீட்கப்படலாம். இதேபோல், NRF2 குறைபாடு எலிகள் ஹிப்போகாம்பஸிலிருந்து காயமடைந்த நியூரான்களில் p62 அளவைக் குறைத்தது [36]. எனவே, NRF2 செயல்படுத்தல் p62 நிலைகளை அதிகரிக்கிறது, இதன் விளைவாக KEAP1 சிதைவு மற்றும் நேர்மறை பின்னூட்ட சுழற்சியில் மேலும் NRF2 உறுதிப்படுத்தலுக்கு ஆதரவாக உள்ளது. NRF2 தூண்டலின் இந்த நியதி அல்லாத பொறிமுறைக்கு மரபணு வெளிப்பாட்டில் மாற்றங்கள் தேவை மற்றும் நீடித்த செல்லுலார் அழுத்தத்திற்கு பொருத்தமான பதிலாக இருக்கலாம்.
சரக்கு அங்கீகார புரதம் NDP52 ஆனது NRF2 ஆல் டிரான்ஸ்கிரிப்ஷனலாக கட்டுப்படுத்தப்பட்டதாகக் காட்டப்பட்டது. NDP52 p62 ஐப் போலவே செயல்படுகிறது, எங்கும் பரவும் புரதங்களை அங்கீகரித்து, LIR டொமைன் மூலம் LC3 உடன் தொடர்பு கொள்கிறது, இதனால் சரக்குகள் லைசோசோம்களில் சிதைக்கப்படுகின்றன. Ndp52 ஊக்குவிப்பாளர் DNA வரிசையில் ஐந்து தூண்டுதல் AREகள் கண்டறியப்பட்டன. NRF2-மத்தியஸ்தம் செய்யப்பட்ட Ndp52 டிரான்ஸ்கிரிப்ஷனுக்கு இன்றியமையாதவை என அவற்றில் மூன்று வெவ்வேறு பிறழ்ந்த கட்டுமானங்கள் மற்றும் ChIP மதிப்பீடுகளுடன் அடையாளம் காணப்பட்டன [113]. குறிப்பு, Nrf52-நாக் அவுட் எலிகளின் ஹிப்போகாம்பஸில் Ndp2 mRNA அளவுகள் குறைக்கப்பட்டன. இந்த வரிசைகளில் ஒன்று NRF2-ஒழுங்குபடுத்தப்பட்ட ARE [36] என ஒரு சுயாதீன ஆய்வில் சரிபார்க்கப்பட்டது.
இருப்பினும், தன்னியக்கத்தின் பண்பேற்றத்தில் NRF2 இன் பங்கு இந்த இரண்டு சரக்கு-அங்கீகார புரதங்களின் தூண்டலுடன் மட்டுப்படுத்தப்படவில்லை. கூடுதல் தன்னியக்கவியல் தொடர்பான மரபணுக்களின் பண்பேற்றத்தில் NRF2 இன் பங்கைப் பற்றிய ஆழமான நுண்ணறிவைப் பெற, எங்கள் குழு NRF1-ஒழுங்குபடுத்தப்பட்ட AREகளை பிணைக்கும் MAFK மற்றும் BACH2 ஆகிய இரண்டு புரதங்களுக்கான குரோமாடின் இம்யூனோபிரெசிபிட்டேஷன் தரவுத்தள ENCODE ஐ திரையிட்டது. JASPAR இன் ஒருமித்த ARE வரிசையிலிருந்து உருவாக்கப்பட்ட ஸ்கிரிப்டைப் பயன்படுத்தி, பல தன்னியக்க மரபணுக்களில் பல தூண்டக்கூடிய AREகளை நாங்கள் அடையாளம் கண்டோம். இந்த வரிசைகளில் 2 வரிசைகள் ஒன்பது தன்னியக்க மரபணுக்களில் NRF2 ஒழுங்குபடுத்தப்பட்ட ARE களாக சரிபார்க்கப்பட்டன, அதன் வெளிப்பாடு Nrf2-நாக் அவுட் எலிகளின் மவுஸ் எம்ப்ரியோ ஃபைப்ரோபிளாஸ்ட்களில் குறைக்கப்பட்டது, ஆனால் NRF2-வெளிப்படுத்தும் லென்டிவைரஸ் மூலம் மீட்டெடுக்க முடியும். தன்னியக்க துவக்கம் (ULK1), சரக்கு அங்கீகாரம் (p62 மற்றும் NDP52), தன்னியக்க உருவாக்கம் (ATG4D, ATG7 மற்றும் GABARAPL1), நீட்டிப்பு (ATG2B மற்றும் ATG5B மற்றும் ATG4B மற்றும் ATG2B மற்றும் ATG36B மற்றும் ATGXNUMXB மற்றும் ATGXNUMXB மற்றும் ATGXNUMXB மற்றும் தன்னியக்க துவக்கம்), தன்னியக்க துவக்கம் (ULKXNUMX), சரக்கு அங்கீகாரம் (pXNUMX மற்றும் NDPXNUMX) உள்ளிட்ட சில மரபணுக்களின் வெளிப்பாட்டை NRFXNUMX செயல்படுத்துகிறது என்பதை எங்கள் ஆய்வு நிரூபித்தது. ), மற்றும் ஆட்டோலிசோசோம் அனுமதி (ATGXNUMXD). இதன் விளைவாக, NRFXNUMX இல்லாதபோது ஹைட்ரஜன் பெராக்சைடுக்கு பதிலளிக்கும் தன்னியக்க ஃப்ளக்ஸ் பலவீனமடைந்தது [XNUMX].
நியூரோடிஜெனரேட்டிவ் கோளாறுகளில் NRF2-மத்தியஸ்த மேக்ரோஆட்டோபாகி ஜீன்களின் வெளிப்பாடு
குறைபாடுள்ள தன்னியக்கமானது பல நரம்பியக்கடத்தல் நோய்களில் முக்கிய பங்கு வகிப்பதாகக் காட்டப்பட்டுள்ளது [114] மற்றும் தன்னியக்கத்தை நீக்குவது எலிகளில் நரம்பியக்கத்திற்கு வழிவகுக்கிறது [115], [116]. Atg7-நாக் அவுட் எலிகள் தன்னியக்கக் குறைபாடு ubiquitin-positive inclution உடல்களில் p62 திரட்சியில் விளைகிறது என்பதை வெளிப்படுத்தியது. இந்த உள்ளடக்கிய உடல்களில் KEAP1 வரிசைப்படுத்தப்பட்டது, இது NRF2 உறுதிப்படுத்தல் மற்றும் இலக்கு மரபணுக்களின் தூண்டலுக்கு வழிவகுத்தது [103]. முக்கியமாக, AD, PD மற்றும் ALS [62] உள்ளிட்ட நரம்பியக்கடத்தல் நோய்களில், எங்கும் பரவும் புரதங்களுடன் p117 இன் அதிகப்படியான குவிப்பு கண்டறியப்பட்டுள்ளது. உண்மையில், AD நோயாளிகளின் உயர் மட்ட APP அல்லது TAU ஐ வெளிப்படுத்தும் நியூரான்கள் p62 மற்றும் நியூக்ளியர் NRF2 ஐ வெளிப்படுத்துகின்றன, இது தன்னியக்கத்தின் மூலம் உள் நரம்புத் தொகுதிகளை சிதைக்கும் முயற்சியை பரிந்துரைக்கிறது [36].
NRF2 குறைபாடு AD இன் சூழலில் புரதத் திரட்டலை மோசமாக்குகிறது. உண்மையில், Nrf2-நாக் அவுட் எலிகளில் பாஸ்போரிலேட்டட் மற்றும் சர்கோசில்-கரையாத TAU இன் அதிகரித்த அளவுகள் காணப்படுகின்றன, இருப்பினும் காட்டு-வகை பின்னணியுடன் ஒப்பிடுகையில் கைனேஸ் அல்லது பாஸ்பேடேஸ் செயல்பாடுகளில் எந்த வித்தியாசமும் கண்டறியப்படவில்லை [113]. முக்கியமாக, NDP52 ஆனது முரைன் நியூரான்களில் TAU உடன் இணை-உள்ளூர்மயமாக்கப்பட்டது மற்றும் பாஸ்போ-TAU மற்றும் NDP52 க்கு இடையேயான நேரடி தொடர்பு எலிகள் மற்றும் AD மாதிரிகள் இரண்டிலும் இணை-இம்யூனோபிரெசிபிட்டேஷன் சோதனைகள் மூலம் காட்டப்பட்டது, இது TAU சிதைவில் அதன் பங்கைச் சுட்டிக்காட்டுகிறது. சுவாரஸ்யமாக, நியூரான்களில் NDP52, p62 அல்லது NRF2 ஐ அமைதிப்படுத்துவது பாஸ்போ-TAU [113], [118] அதிகரித்தது. மேலும், NRF1 இல்லாதபோது, APP/PS9?E2 எலிகளின் ஹிப்போகாம்பஸில் அதிகரித்த அகநியூரோனல் APP திரட்டுகள் கண்டறியப்பட்டன. இது மாற்றப்பட்ட தன்னியக்க குறிப்பான்களுடன் தொடர்புடையது, இதில் அதிகரித்த பாஸ்போ-எம்டிஓஆர்/எம்டிஓஆர் மற்றும் பாஸ்போ-பி70எஸ்6கே/பி70எஸ்6கே விகிதங்கள் (தன்னியக்கத் தடுப்பைக் குறிக்கிறது), முன்-கேதெப்சின் டி மற்றும் அதிக எண்ணிக்கையிலான மல்டிவிசிகுலர் உடல்கள் [119]. எலிகள் இணைந்து வெளிப்படுத்தும் மனித APP (V717I) மற்றும் TAU (P301L), NRF2 குறைபாடானது கரையாத பகுதியிலுள்ள மொத்த மற்றும் பாஸ்போ-TAU அளவை அதிகரிக்க வழிவகுத்தது. ATG62 மற்றும் GABARAPL52. அடாப்டர் புரதம் p1 மற்றும் APP அல்லது TAU ஆகியவற்றுக்கு இடையேயான இணை-உள்ளூர்மயமாக்கல் NRF5 இல்லாத நிலையில் குறைக்கப்பட்டது [1]. ஒட்டுமொத்தமாக, இந்த முடிவுகள் நரம்பியல் தன்னியக்கத்தில் NRF62 இன் முக்கியத்துவத்தை எடுத்துக்காட்டுகின்றன.
வெவ்வேறு டிரான்ஸ்கிரிப்ஷன் காரணிகள் புரோட்டியோஸ்டாசிஸை மாடுலேட் செய்ய ஒருங்கிணைந்து செயல்படுகின்றன
நிலையான நிலையின் கீழ், புரோட்டீன்-புரத இடைவினைகள் மற்றும் விரைவான பதிலைப் பெறும் மொழிபெயர்ப்புக்குப் பிந்தைய மாற்றங்கள் மூலம் புரோட்டியோஸ்டாஸிஸ் கட்டுப்படுத்தப்படுகிறது. இருப்பினும், செல்லுலார் தழுவலுக்கு UPR, UPS மற்றும் தன்னியக்க மரபணுக்களின் டிரான்ஸ்கிரிப்ஷனல் ஒழுங்குமுறை தேவைப்படுகிறது. ஆக்ஸிஜனேற்றம் மற்றும் புரோட்டியோடாக்ஸிக் அழுத்தம் உள்ளிட்ட குறைந்த தர நச்சு அவமதிப்புகளுக்கு நரம்பு செல்கள் தொடர்ந்து சமர்ப்பிக்கப்படுவதைக் கருத்தில் கொண்டு, டிரான்ஸ்கிரிப்ஷனல் மாடுலேஷன் மூலம் தூண்டப்பட்ட புரோட்டியோஸ்டாசிஸின் வலுவூட்டல் மூளை சிதைவைத் தடுக்க உதவும்.
UPR ஐப் பொறுத்தவரை, மூன்று கைகளில் ஒவ்வொன்றின் செயல்பாட்டின் விளைவாக, சில மரபணுக்களின் டிரான்ஸ்கிரிப்ஷனல் தூண்டல் இறுதியில் ஏற்படும் ([43] இல் மதிப்பாய்வு செய்யப்பட்டது). எடுத்துக்காட்டாக, ATF6-பெறப்பட்ட துண்டு (ATF6f) ER- அழுத்த மறுமொழி கூறுகளுடன் (ERSE) பிணைக்கிறது மற்றும் XBPI, BIP மற்றும் CHOP உட்பட பல மரபணுக்களின் வெளிப்பாட்டைத் தூண்டுகிறது. கூடுதலாக, PERK சிக்னலிங் டிரான்ஸ்கிரிப்ஷன் காரணி ATF4 ஐ செயல்படுத்த வழிவகுக்கிறது, இது பல UPR தொடர்பான மரபணுக்களின் வெளிப்பாட்டைக் கட்டுப்படுத்துகிறது மற்றும் NRF2 இலக்கு மரபணுக்கள் Hmox1 மற்றும் p62 உட்பட சிலவற்றைக் கட்டுப்படுத்துகிறது. இறுதியாக, IRE1 செயல்படுத்துதலானது செயலில் உள்ள டிரான்ஸ்கிரிப்ஷன் காரணி, பிளவுபட்ட XBP1 (XBP1s) உருவாக்கத்தில் விளைகிறது, இது புரத மடிப்புகளில் ஈடுபடும் மரபணுக்களின் குறியாக்கப் புரதங்களின் டிரான்ஸ்கிரிப்ஷனைக் கட்டுப்படுத்துகிறது.
மறுபுறம், NRF1 மூளையில் புரோட்டீசோமால் மரபணு வெளிப்பாட்டிற்கு அவசியமானது எனக் காட்டப்பட்டது, ஏனெனில் Nrf1-நாக் அவுட் எலிகள் 20S மையத்தின் பல்வேறு துணைப்பிரிவுகளை குறியாக்கம் செய்யும் மரபணுக்களின் குறைக்கப்பட்ட வெளிப்பாட்டை வெளிப்படுத்தின. ]. NRF19 மற்றும் NRF90 இரண்டும் அதன் இலக்கு மரபணுக்களின் ஊக்குவிப்புப் பகுதிகளில் ARE வரிசைகளுடன் பிணைக்கப்படுகின்றன, அவை டிரான்ஸ்கிரிப்ஷனல் செயல்பாடுகளை ஒன்றுடன் ஒன்று சேர்க்கின்றன, இருப்பினும் அவை அவற்றின் ஒழுங்குமுறை வழிமுறைகள் மற்றும் செல்லுலார் உள்ளூர்மயமாக்கலில் வேறுபடுகின்றன [1].
ஃபோர்க்ஹெட் பாக்ஸ் O (FOXO) குடும்பத்தின் டிரான்ஸ்கிரிப்ஷன் காரணிகள் பல தன்னியக்கவியல் தொடர்பான மரபணுக்களின் வெளிப்பாட்டைக் கட்டுப்படுத்துகின்றன. NRF2 உடன் நிகழ்வதைப் போலவே, FOXO உறுப்பினர்களின் செயல்பாட்டை ஒழுங்குபடுத்தும் பல அடுக்குகள் உள்ளன, அவை ஊட்டச்சத்து அல்லது ஆக்ஸிஜனேற்ற அழுத்தத்தின் மீது தூண்டப்படலாம் [121]. இறுதியாக, டிரான்ஸ்கிரிப்ஷன் காரணி TFEB, லைசோசோமால் பயோஜெனீசிஸின் முதன்மை சீராக்கியாகக் கருதப்படுகிறது, ஊட்டச்சத்து அழுத்த நிலைமைகளின் கீழ் தன்னியக்கத்தைக் கட்டுப்படுத்துவதில் முக்கிய பங்கு வகிக்கிறது. எனவே, mTORC1 இன் தடுப்பானது TFEB இன் அணுக்கரு இடமாற்றம் மற்றும் தன்னியக்க மரபணுக்களின் வெளிப்பாட்டின் தூண்டுதலுக்கு வழிவகுக்கிறது [122].
ஒட்டுமொத்தமாக, இந்த இயந்திரங்களின் வெவ்வேறு டிரான்ஸ்கிரிப்ஷனல் ரெகுலேட்டர்களின் இருப்பு வெவ்வேறு சூழ்நிலைகளில் புரோட்டியோஸ்டாசிஸை உறுதிப்படுத்தக்கூடிய க்ரோஸ்டாக் மற்றும் ஓரளவு தேவையற்ற வழிமுறைகளையும் பரிந்துரைக்கிறது. அதன்படி, அதிக அளவு ஆக்ஸிஜனேற்ற அழுத்தத்தை ஆதரிக்கும் திசுக்களில் NRF2 பொருத்தமான பங்கைக் கொண்டிருக்கலாம். எடுத்துக்காட்டாக, ஆக்ஸிஜனேற்ற அழுத்தத்தால் தூண்டப்பட்ட NRF2, பட்டினி நிலையில் TFEB க்கு கண்டறியப்பட்டதைப் போலவே, டிரான்ஸ்கிரிப்ஷனலாக தன்னியக்கத்தைக் கட்டுப்படுத்த ஊட்டச்சத்து நிறைந்த நிலைமைகளின் கீழ் செயல்படலாம். மேலும், மூளை பெரும்பாலும் ஊட்டச்சத்து நிறைந்த நிலைமைகளின் கீழ் செயல்படுகிறது, நியூரான்களில் தன்னியக்கத்தை செயல்படுத்துவதற்கு NRF2 ஒரு பொருத்தமான வழிமுறையாக உள்ளது.
புரோட்டினோபதிகளில் NRF2க்கான நம்பிக்கைக்குரிய சிகிச்சை சாத்தியம்
கடந்த சில ஆண்டுகளில், UPR, UPS மற்றும் NRF2 செயல்பாட்டில் தன்னியக்கத்தின் ஒழுங்குமுறை பாத்திரங்கள் மற்றும் இந்த மூன்று அமைப்புகளின் கூறுகளின் பரஸ்பர NRF2-மத்தியஸ்த டிரான்ஸ்கிரிப்ஷன் ஆகியவற்றின் அறிவில் பெரும் முன்னேற்றம் ஏற்பட்டுள்ளது. எனவே, நியூரோடிஜெனரேடிவ் நோய்களில் புரோட்டீன் அனுமதியின் முக்கியமான சீராக்கியாக NRF2 சுரண்டலின் அடிப்படையில் புதிய சிகிச்சை சாத்தியங்கள் எழலாம்.
இருப்பினும், மூளையில் NRF2 அளவை அதிகரிப்பது பயனுள்ளதா அல்லது தீங்கு விளைவிப்பதா என்பது ஒரு முக்கியமான கேள்வி. NFE2L2 மரபணு மிகவும் பாலிமார்ஃபிக் மற்றும் அதன் ஊக்குவிப்பு ஒழுங்குமுறை பகுதியில் காணப்படும் சில ஒற்றை நியூக்ளியோடைடு பாலிமார்பிஸங்கள் மக்கள்தொகை மட்டத்தில் மரபணு வெளிப்பாட்டின் "உடலியல்" மாறுபாட்டின் வரம்பை வழங்கலாம் மற்றும் சில ஹாப்லோடைப்களை வழங்குவதால், தொற்றுநோயியல் தரவுகளின் பகுப்பாய்வு ஒரு பகுதியளவு பதிலை அளிக்கலாம். AD, PD அல்லது ALS [123] இன் குறைவான ஆபத்து மற்றும்/அல்லது தாமதமான தொடக்கத்துடன் தொடர்புடையது. மேலும், ஹேய்ஸ் மற்றும் சகாக்களால் [124] விவாதிக்கப்பட்டபடி, NRF2 விளைவு U-வடிவ பதிலைக் கொண்டிருக்கலாம், அதாவது மிகக் குறைந்த NRF2 அளவுகள் சைட்டோபுரோடெக்ஷனை இழக்க நேரிடலாம் மற்றும் அழுத்தங்களுக்கு அதிக உணர்திறன் ஏற்படலாம், அதே நேரத்தில் அதிக அளவு NRF2 ஹோமியோஸ்ட்டிக் சமநிலையைத் தொந்தரவு செய்யலாம் ஒரு குறைக்கும் சூழ்நிலை (குறைப்பு அழுத்தம்), இது புரதம் தவறாக மடித்தல் மற்றும் திரட்டுதல் ஆகியவற்றிற்கு சாதகமாக இருக்கும். மூளையில் உள்ள குறைந்த NRF2 அளவுகள், நோயியல் நிலைமைகளின் கீழ் ஒரு நன்மையை அடைய ஒரு சிறிய கட்டுப்பாடு போதுமானதாக இருக்கலாம் என்ற கருத்தை ஆதரிக்கிறது. உண்மையில், புரோட்டீன் அனுமதியின் மருந்தியல் NRF2-மத்தியஸ்த செயல்பாட்டின் பாதுகாப்புப் பங்கு வெவ்வேறு நரம்பியக்க உயிரணு கலாச்சாரம் மற்றும் விவோ மாதிரிகளில் காட்டப்பட்டுள்ளது.
SFN என்பது ஒரு மருந்தியல் NRF2 ஆக்டிவேட்டர் ஆகும், இது புரோட்டீசோமால் மற்றும் தன்னியக்க மரபணு வெளிப்பாட்டைத் தூண்டுவதற்கு நிரூபிக்கப்பட்டது [95], [36]. சுவாரஸ்யமாக, SFN பாஸ்போரிலேட்டட் TAU இன் அளவைக் குறைத்தது மற்றும் பெக்லின்-1 மற்றும் LC3-II ஐ அதிகரித்தது என்பதை ஜோ மற்றும் சகாக்கள் நிரூபித்தனர், NRF2 செயல்படுத்தல் தன்னியக்கத்தின் மூலம் இந்த நச்சு புரதத்தின் சிதைவை எளிதாக்கலாம் என்று பரிந்துரைக்கிறது [113]. மேலும், mHtt இன் சிதைவு SFN உடன் மேம்படுத்தப்பட்டது, மேலும் இது MG132 ஐப் பயன்படுத்தி மாற்றியமைக்கப்பட்டது, இது இந்த நச்சு புரதத்தின் புரோட்டீசோமால் சிதைவைக் குறிக்கிறது [95]. கரிம ஃபிளாவனாய்டு ஃபிசெட்டின் மூலம் பாஸ்போ மற்றும் கரையாத-TAU இன் தன்னியக்க-மத்தியஸ்த சிதைவு தெரிவிக்கப்பட்டது. இந்த கலவையானது TFEB மற்றும் NRF2 இரண்டையும் அதன் இலக்கு மரபணுக்களுடன் ஒரே நேரத்தில் செயல்படுத்துதல் மற்றும் அணுக்கரு இடமாற்றத்தை ஊக்குவிப்பதன் மூலம் தன்னியக்கத்தைத் தூண்ட முடிந்தது. இந்த பதில் TFEB அல்லது NRF2 அமைதிப்படுத்தல் மூலம் தடுக்கப்பட்டது [125]. பாட் மற்றும் சகாக்கள் ஒரே நேரத்தில் NRF2, NRF1 மற்றும் HSF1 ஆக்டிவேட்டரின் நன்மையான விளைவுகளை ஸ்பைனல் மற்றும் பல்பார் மஸ்குலர் அட்ராபியில் புரோட்டீன் நச்சுத்தன்மையின் மீது தெரிவித்துள்ளனர் மல்டிபிள் ஸ்களீரோசிஸ் [126], [2] சிகிச்சைக்காக NRF12 தூண்டி டைமிதில் ஃபுமரேட்டின் (DMF) வாய்வழி உருவாக்கமான BG-2 இன் ஒப்புதலுடன் நியூரோடிஜெனரேடிவ் கோளாறுகளுக்கான சிகிச்சைக்கான NRF127 செயல்படுத்தும் சாத்தியம் நிரூபிக்கப்பட்டுள்ளது. ஒரு வலுவான அழற்சி கூறு கொண்ட தன்னுடல் தாக்க நோய்களுடன் DMF இன் வெற்றி, இந்த மருந்தை மாற்றியமைப்பதன் மூலம் நரம்பியக்கடத்தல் நோய்கள் பயனடையக்கூடும் என்று கூறுகிறது. PD இன் ?-சினுக்ளினோபதி மாதிரியின் சமீபத்திய முன்கூட்டிய ஆய்வில், DMF அதன் தன்னியக்கத் தூண்டுதலின் ஒரு பகுதியாக நியூரோபிராக்டிவ் எனக் காட்டப்பட்டது [128]. நியூரோடிஜெனரேஷனில் NRF129 இன் நன்மை பயக்கும் விளைவுகளைப் புகாரளிக்கும் ஆய்வுகள், ஆனால் புரோட்டீன் கிளியரன்ஸ் மீதான அதன் விளைவை மையமாகக் கொண்டிருக்கவில்லை (ஒரு விரிவான மதிப்பாய்விற்கு, [2] பார்க்கவும்). ஆக்ஸிஜனேற்ற அழுத்தம், நியூரோஇன்ஃப்ளமேஷன் அல்லது மைட்டோகாண்ட்ரியல் செயலிழப்பு உள்ளிட்ட NRF7 இல் ஒரே நேரத்தில் ஒரே நேரத்தில் இலக்கு வைக்கக்கூடிய பல சேதப்படுத்தும் செயல்முறைகளை இது எடுத்துக்காட்டுவதால் இது மிகவும் பொருத்தமானது. இருப்பினும், மூளையில் உள்ள நச்சு புரதங்களின் சிதைவை எளிதாக்குவதற்கு NRF2 இன் மருந்தியல் செயலாக்கம் சரியான உத்தியாக இருக்குமா என்பதை நிச்சயமாக தீர்மானிக்க எதிர்கால வேலை தேவைப்படும்.
முன்பு விளக்கியது போல், GSK-3 அதிகப்படுத்தப்பட்டதா? நியூரோடிஜெனரேடிவ் நோய்களில் செயல்பாடு பதிவாகியுள்ளது மற்றும் அதன் விளைவாக NRF2 குறைப்பு தீங்கு விளைவிக்கும் விளைவுகளுக்கு ஓரளவு காரணமாக இருக்கலாம் என்று ஊகிக்கப்பட்டது. இந்த நோயியல் நிலைமைகளின் கீழ், GSK-3 தடுப்பான்கள் NRF2 அளவுகள் மற்றும் புரோட்டியோஸ்டாசிஸை அதிகரிக்க ஒத்துழைக்கலாம். GSK-3 இன்ஹிபிட்டர்களின் நன்மையான விளைவுகள் நியூரோடிஜெனரேஷனின் வெவ்வேறு மாதிரிகளில் பதிவாகியுள்ளன, மேலும் சுவாரஸ்யமாக, GSK-3 அடக்குமுறை நச்சு புரதங்களின் அளவைக் குறைப்பதாகக் காட்டப்பட்டது [130], [131], [132], [133]. GSK-3 தடுப்பு மற்றும் புரோட்டியோஸ்டாசிஸை ஊக்குவிக்கும் மரபணுக்களின் NRF2-டிரான்ஸ்கிரிப்ஷனல் ஒழுங்குமுறை ஆகியவற்றுக்கு இடையே நேரடித் தொடர்புகள் எதுவும் இதுவரை காணப்படவில்லை என்றாலும், GSK-3 செயல்பாட்டின் கீழ்-கட்டுப்பாடு NRF2 அளவுகளை அதிகரிக்கும் என்று ஊகிக்க நியாயமானது, இது இறுதியில் வலுவூட்டப்படும். புரோட்டியோஸ்டாஸிஸ்.
NRF2 இன் டிரான்ஸ்கிரிப்ஷனல் செயல்பாடு மற்றும் புரோட்டியோஸ்டாசிஸைப் பராமரிப்பதற்கான செல்லுலார் திறன் வயதுக்கு ஏற்ப குறைகிறது, இது நியூரோடிஜெனரேடிவ் நோய்களின் வளர்ச்சிக்கான முக்கிய ஆபத்து காரணியாகும். NRF2 இன் வலுவூட்டல் மற்றும் அதன் விளைவாக, புரோட்டியோஸ்டாசிஸ், குறைந்தபட்சம், புரதத் திரட்டுகள் மற்றும் நரம்பணு சிதைவைத் தாமதப்படுத்தும் என்று நினைப்பது நியாயமானது. உண்மையில், 18?-கிளைசிர்ஹெட்டினிக் அமிலம் (18?-GA) ட்ரைடர்பெனாய்டு கொண்ட மனித முதிர்ந்த ஃபைப்ரோபிளாஸ்ட்களின் சிகிச்சையானது NRF2 செயல்படுத்தலை ஊக்குவித்தது, இது புரோட்டீசோம் தூண்டுதலுக்கும் மேம்பட்ட ஆயுட்காலத்திற்கும் வழிவகுத்தது. NRF2 இன் மருந்தியல் செயலாக்கம் பிற்பகுதியில் கூட சாத்தியமாகும் என்று இந்த ஆய்வு தெரிவிக்கிறது [86]. மேலும், இந்த கலவை SKN-1 மற்றும் C.elegans இல் புரோட்டீசோம் செயல்படுத்தலை மத்தியஸ்தம் செய்தது, தொடர்புடைய நூற்புழு மாதிரிகள் [134] AD முன்னேற்றத்தில் நன்மை பயக்கும்.
எல்லாவற்றையும் கருத்தில் கொண்டால், புரோட்டியோஸ்டாஸிஸ் தொடர்பான மரபணுக்களின் NRF2-மத்தியஸ்த தூண்டல் வெவ்வேறு புரோட்டினோபதிகளில் நன்மை பயக்கும்.
புற்றுநோய், இறப்பு, முதுமை, மூளை மற்றும் நடத்தை, இதய நோய் மற்றும் பலவற்றில் சல்போராபேன் மற்றும் அதன் விளைவுகள்
ஐசோதியோசயனேட்டுகள் உங்கள் உணவில் நீங்கள் பெறக்கூடிய மிக முக்கியமான தாவர கலவைகள் ஆகும். இந்த வீடியோவில் நான் அவர்களுக்காக இதுவரை செய்யப்படாத மிக விரிவான வழக்கை உருவாக்குகிறேன். குறுகிய கவனம்? கீழே உள்ள நேரப் புள்ளிகளில் ஒன்றைக் கிளிக் செய்வதன் மூலம் உங்களுக்குப் பிடித்த தலைப்புக்குச் செல்லவும். முழு காலவரிசை கீழே.
முக்கிய பிரிவுகள்:
00:01:14 - புற்றுநோய் மற்றும் இறப்பு
00:19:04 - முதுமை
00:26:30 - மூளை மற்றும் நடத்தை
00:38:06 - இறுதி மறுபரிசீலனை
00:40:27 - டோஸ்
முழு காலவரிசை:
00:00:34 – வீடியோவின் முக்கிய மையமான சல்போராபேன் அறிமுகம்.
00:01:14 - க்ரூசிஃபெரஸ் காய்கறி நுகர்வு மற்றும் அனைத்து காரணங்களின் இறப்பு குறைப்பு.
00:02:12 - புரோஸ்டேட் புற்றுநோய் ஆபத்து.
00:02:23 - சிறுநீர்ப்பை புற்றுநோய் ஆபத்து.
00:02:34 - புகைப்பிடிப்பவர்களுக்கு நுரையீரல் புற்றுநோய் ஆபத்து.
00:02:48 - மார்பக புற்றுநோய் ஆபத்து.
00:03:13 - அனுமானம்: உங்களுக்கு ஏற்கனவே புற்றுநோய் இருந்தால் என்ன செய்வது? (இடையிடல்)
00:03:35 – புற்றுநோய் மற்றும் இறப்பு தொடர்பான தரவுகளை இயக்கும் நம்பத்தகுந்த வழிமுறை.
00:04:38 - சல்போராபேன் மற்றும் புற்றுநோய்.
00:05:32 - எலிகளில் சிறுநீர்ப்பை கட்டி வளர்ச்சியில் ப்ரோக்கோலி முளை சாறு வலுவான விளைவைக் காட்டும் விலங்கு சான்றுகள்.
00:06:06 - புரோஸ்டேட் புற்றுநோயாளிகளில் சல்ஃபோராபேன் நேரடியாக நிரப்புவதன் விளைவு.
00:07:09 - உண்மையான மார்பக திசுக்களில் ஐசோதியோசயனேட் வளர்சிதை மாற்றங்களின் உயிர் குவிப்பு.
00:08:32 - மார்பக புற்றுநோய் ஸ்டெம் செல்களைத் தடுப்பது.
00:08:53 - வரலாற்றுப் பாடம்: பண்டைய ரோமில் கூட பிராசிகாக்கள் ஆரோக்கிய பண்புகளைக் கொண்டிருப்பதாக நிறுவப்பட்டது.
00:09:16 – சல்போராபேன் புற்றுநோய் வெளியேற்றத்தை மேம்படுத்தும் திறன் (பென்சீன், அக்ரோலின்).
00:09:51 - ஆன்டிஆக்ஸிடன்ட் மறுமொழி கூறுகள் வழியாக மரபணு மாற்றமாக NRF2.
00:10:10 – NRF2 ஆக்டிவேஷன் குளுதாதயோன்-எஸ்-கான்ஜுகேட்ஸ் வழியாக புற்றுநோயை வெளியேற்றுவதை எவ்வாறு மேம்படுத்துகிறது.
00:10:34 - பிரஸ்ஸல்ஸ் முளைகள் குளுதாதயோன்-எஸ்-டிரான்ஸ்ஃபெரேஸை அதிகரிக்கின்றன மற்றும் டிஎன்ஏ பாதிப்பைக் குறைக்கின்றன.
00:11:20 - ப்ரோக்கோலி முளை பானம் பென்சீன் வெளியேற்றத்தை 61% அதிகரிக்கிறது.
00:13:31 - ப்ரோக்கோலி ஸ்ப்ரூட் ஹோமோஜெனேட் மேல் சுவாசப்பாதையில் ஆக்ஸிஜனேற்ற நொதிகளை அதிகரிக்கிறது.
00:15:45 - சிலுவை காய்கறி நுகர்வு மற்றும் இதய நோய் இறப்பு.
00:16:55 - ப்ரோக்கோலி ஸ்ப்ரூட் பவுடர் இரத்த கொழுப்பு மற்றும் ஒட்டுமொத்த இதய நோய் அபாயத்தை மேம்படுத்துகிறது வகை 2 நீரிழிவு நோயாளிகள்.
00:19:04 - வயதான பிரிவின் ஆரம்பம்.
00:19:21 - சல்போராபேன்-செறிவூட்டப்பட்ட உணவு வண்டுகளின் ஆயுட்காலம் 15 முதல் 30% வரை அதிகரிக்கிறது (சில நிபந்தனைகளில்).
00:20:34 - நீண்ட ஆயுளுக்கு குறைந்த வீக்கத்தின் முக்கியத்துவம்.
00:22:05 - குரூசிஃபெரஸ் காய்கறிகள் மற்றும் ப்ரோக்கோலி ஸ்ப்ரூட் பவுடர் மனிதர்களில் பல்வேறு வகையான அழற்சி குறிப்பான்களைக் குறைப்பதாகத் தெரிகிறது.
00:23:40 – மிட்-வீடியோ ரீகேப்: புற்றுநோய், வயதான பிரிவுகள்
00:24:14 - முதுமையில் சல்போராபேன் தகவமைப்பு நோயெதிர்ப்புச் செயல்பாட்டை மேம்படுத்தலாம் என்று சுட்டி ஆய்வுகள் தெரிவிக்கின்றன.
00:25:18 – சல்ஃபோராபேன், வழுக்கை மவுஸ் மாதிரியில் முடி வளர்ச்சியை மேம்படுத்தியது. படம் 00:26:10.
00:28:17 - மனச்சோர்வு விவாதத்தின் தொடக்கம் (நம்பத்தகுந்த வழிமுறை மற்றும் ஆய்வுகள்).
00:31:21 – அழுத்தத்தால் தூண்டப்பட்ட மனச்சோர்வின் 10 வெவ்வேறு மாதிரிகளைப் பயன்படுத்தி மவுஸ் ஆய்வு, ஃப்ளூக்ஸெடின் (ப்ரோசாக்) போலவே சல்ஃபோராபேன் பயனுள்ளதாக இருக்கும்.
00:32:00 – எலிகளில் குளுக்கோராபனின் நேரடியாக உட்கொள்வது சமூக தோல்வி மன அழுத்த மாதிரியிலிருந்து மனச்சோர்வைத் தடுப்பதில் இதேபோல் பயனுள்ளதாக இருக்கும் என்று ஆய்வு காட்டுகிறது.
00:33:01 - நியூரோடிஜெனரேஷன் பிரிவின் ஆரம்பம்.
00:33:30 – சல்போராபேன் மற்றும் அல்சைமர் நோய்.
00:33:44 – சல்போராபேன் மற்றும் பார்கின்சன் நோய்.
00:33:51 – சல்போராபேன் மற்றும் ஹங்டிங்டன் நோய்.
00:34:13 - சல்ஃபோராபேன் வெப்ப அதிர்ச்சி புரதங்களை அதிகரிக்கிறது.
00:34:43 - அதிர்ச்சிகரமான மூளை காயம் பிரிவின் ஆரம்பம்.
00:35:55 - சல்ஃபோராபேன் மற்றும் நியூரானல் பிளாஸ்டிசிட்டி.
00:36:32 – எலிகளில் டைப் II நீரிழிவு மாதிரியில் சல்போராபேன் கற்றலை மேம்படுத்துகிறது.
00:37:19 - சல்போராபேன் மற்றும் டுச்சேன் தசைநார் சிதைவு.
00:37:44 - தசை செயற்கைக்கோள் செல்களில் Myostatin தடுப்பு (விட்ரோவில்).
00:38:06 – லேட்-வீடியோ மறுபரிசீலனை: இறப்பு மற்றும் புற்றுநோய், டிஎன்ஏ சேதம், ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் வீக்கம், பென்சீன் வெளியேற்றம், இருதய நோய், வகை II நீரிழிவு, மூளையில் விளைவுகள் (மன அழுத்தம், மன இறுக்கம், ஸ்கிசோஃப்ரினியா, நியூரோடிஜெனரேஷன்), NRF2 பாதை.
00:40:27 - ப்ரோக்கோலி முளைகள் அல்லது சல்ஃபோராபேன் அளவைக் கண்டறிவது பற்றிய எண்ணங்கள்.
00:41:01 - வீட்டில் முளைப்பது பற்றிய நிகழ்வுகள்.
00:43:14 - சமையல் வெப்பநிலை மற்றும் சல்ஃபோராபேன் செயல்பாடு.
00:43:45 - குளுகோராபனினில் இருந்து சல்போராபேன் குடல் பாக்டீரியாவை மாற்றுகிறது.
00:44:24 - காய்கறிகளிலிருந்து செயலில் உள்ள மைரோசினேஸுடன் இணைந்தால் சப்ளிமெண்ட்ஸ் சிறப்பாகச் செயல்படும்.
00:44:56 - சமையல் நுட்பங்கள் மற்றும் சிலுவை காய்கறிகள்.
00:46:06 - ஐசோதியோசயனேட்டுகள் கோயிட்ரோஜன்களாக.
நியூக்ளியர் காரணி எரித்ராய்டு-பெறப்பட்ட 2 (NF-E2)-தொடர்புடைய காரணி 2, இல்லையெனில் Nrf2 என அழைக்கப்படுகிறது, இது ஒரு டிரான்ஸ்கிரிப்ஷன் காரணியாகும், இது பல்வேறு ஆக்ஸிஜனேற்ற மற்றும் நச்சுத்தன்மையற்ற நொதிகளின் வெளிப்பாட்டைக் கட்டுப்படுத்துகிறது. ஆக்ஸிஜனேற்ற அழுத்தத்தைக் கட்டுப்படுத்துவதில் அதன் பங்கை ஆராய்ச்சி ஆய்வுகள் நிரூபித்துள்ளன. அல்சைமர் நோய் மற்றும் பார்கின்சன் நோய் போன்ற பெரும்பாலான நரம்பியக்கடத்தல் நோய்கள், ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் நாள்பட்ட அழற்சியால் வகைப்படுத்தப்படுகின்றன, பொதுவான இலக்குகள் Nrf2 சிகிச்சை அணுகுமுறைகள். டாக்டர். அலெக்ஸ் ஜிமினெஸ் DC, CCST இன்சைட்
இறுதியான குறிப்புகள்
டிரான்ஸ்கிரிப்ஷன் காரணி NRF2 UPR, UPS மற்றும் ஆட்டோபேஜி (படம் 4) ஆகியவற்றில் ஏற்படும் மாற்றங்களை உணர்ந்து மாற்றியமைப்பதன் மூலம் ஒரு புரோட்டியோஸ்டேடிக் பதிலைத் திட்டமிடுகிறது. இதன் விளைவாக, NRF2 இன் பற்றாக்குறை புரோட்டினோபதியை மோசமாக்குவதாகக் காட்டப்பட்டுள்ளது, இது உகந்த புரத அனுமதிக்கு NRF2 அவசியம் என்று பரிந்துரைக்கிறது. அனைத்தும் சேர்ந்து, புரோட்டினோபதிகளுக்கு NRF2 ஒரு சுவாரஸ்யமான சிகிச்சை இலக்காக இருக்கலாம் என்று நாம் ஊகிக்க முடியும்.
படம் 4 NRF2, புரோட்டியோடாக்ஸிக்-பெறப்பட்ட அவசரகால சமிக்ஞைகளை ஒரு பாதுகாப்பு டிரான்ஸ்கிரிப்ஷனல் பதிலுடன் இணைக்கும் மையமாக உள்ளது. விரிக்கப்பட்ட/தவறாக மடிக்கப்பட்ட புரதங்களின் திரட்சியானது ER இல் விரிக்கப்பட்ட புரத மறுமொழியை (UPR) செயல்படுத்த வழிவகுக்கும். PERK அல்லது MAPK ஐச் செயல்படுத்துவது ER-குடியிருப்பு Gpx8 இன் டிரான்ஸ்கிரிப்ஷனல் தூண்டலுக்கும் மற்றும் GSH அளவைக் கட்டுப்படுத்தும் பல நொதிகளுக்கும் வழிவகுக்கும், இது சரியான புரத மடிப்புக்கு முக்கியமானது. புரோட்டீசோம் செயல்பாட்டை (யுபிஎஸ்) புரதத் திரட்டுகள் தடுக்கின்றன, ஒருவேளை NRF2 சிதைவைத் தவிர்க்கலாம். NRF2 குறிப்பாக Psma3, Psma6, Psmb1, Psmb5 மற்றும் Pomp மரபணுக்களின் படியெடுத்தலை மாற்றியமைப்பதாகக் காட்டப்பட்டுள்ளது. D2T க்கு பதிலளிக்கும் விதமாக NRF3-சார்ந்த முறையில் வேறு பல துணைக்குழுக்கள் கட்டுப்படுத்தப்பட்டன, ஒருவேளை NRF2 ஆல் கட்டுப்படுத்தப்படும் புரோட்டீசோம் துணைக்குழுக்களின் பட்டியலை பெரிதாக்கலாம். தன்னியக்கமானது புரதத் திரட்டுகளின் சிதைவுக்கான முக்கிய பாதையாகும். தன்னியக்கவியல் NRF2 ஐ ஒழுங்குபடுத்துகிறது, இந்த சிதைவு பாதையை p2, Ndp62, Ulk52, Atg1b, Atg2c, Atg4, Atg5 மற்றும் Gabarapl7 ஆகியவற்றின் NRF1 டிரான்ஸ்கிரிப்ஷனல் தூண்டலுடன் இணைக்கிறது.
மேலே உள்ள கட்டுரையின் படி, நரம்பியக்கடத்தல் நோய்களின் அறிகுறிகளை பல்வேறு சிகிச்சை விருப்பங்கள் மூலம் சிகிச்சையளிக்க முடியும், ஆராய்ச்சி ஆய்வுகள் Nrf2 செயல்படுத்தல் ஒரு பயனுள்ள சிகிச்சை அணுகுமுறையாக இருக்கும் என்பதை நிரூபித்துள்ளது. ஏனெனில் Nrf2 ஆக்டிவேட்டர்கள் நோயின் பரந்த வழிமுறைகளை குறிவைக்கின்றன, அனைத்து நரம்பியக்கடத்தல் நோய்களும் Nrf2 டிரான்ஸ்கிரிப்ஷன் காரணியைப் பயன்படுத்துவதன் மூலம் பயனடையலாம். Nrf2 இன் கண்டுபிடிப்புகள் நியூரோடிஜெனரேட்டிவ் நோய்களுக்கான சிகிச்சையில் புரட்சியை ஏற்படுத்தியுள்ளன. எங்கள் தகவலின் நோக்கம் உடலியக்க மற்றும் முதுகெலும்பு சுகாதார பிரச்சினைகள் மட்டுமே. விஷயத்தைப் பற்றி விவாதிக்க, தயவு செய்து டாக்டர் ஜிமினெஸைக் கேட்கவும் அல்லது எங்களை இங்கே தொடர்பு கொள்ளவும்915-850-0900.
கூடுதல் தலைப்பு விவாதம்: அறுவை சிகிச்சை இல்லாமல் முழங்கால் வலியை நீக்குதல்
முழங்கால் வலி என்பது நன்கு அறியப்பட்ட அறிகுறியாகும், இது பல்வேறு முழங்கால் காயங்கள் மற்றும்/அல்லது நிலைமைகள் காரணமாக ஏற்படலாம்.விளையாட்டு காயங்கள். முழங்கால் மனித உடலில் மிகவும் சிக்கலான மூட்டுகளில் ஒன்றாகும், ஏனெனில் இது நான்கு எலும்புகள், நான்கு தசைநார்கள், பல்வேறு தசைநாண்கள், இரண்டு மெனிசிஸ் மற்றும் குருத்தெலும்பு ஆகியவற்றின் குறுக்குவெட்டுகளால் ஆனது. அமெரிக்கன் அகாடமி ஆஃப் ஃபேமிலி பிசிஷியன்களின் கூற்றுப்படி, முழங்கால் வலிக்கான பொதுவான காரணங்களில் பட்டேலர் சப்லக்சேஷன், பட்டேலர் டெண்டினிடிஸ் அல்லது ஜம்பர்ஸ் முழங்கால் மற்றும் ஓஸ்குட்-ஸ்க்லாட்டர் நோய் ஆகியவை அடங்கும். முழங்கால் வலி 60 வயதுக்கு மேற்பட்டவர்களுக்கு ஏற்பட வாய்ப்புகள் இருந்தாலும், குழந்தைகள் மற்றும் இளம்பருவத்தினருக்கும் முழங்கால் வலி ஏற்படலாம். முழங்கால் வலியை RICE முறைகளைப் பின்பற்றி வீட்டிலேயே சிகிச்சை செய்யலாம், இருப்பினும், கடுமையான முழங்கால் காயங்களுக்கு உடலியக்க சிகிச்சை உட்பட உடனடி மருத்துவ கவனிப்பு தேவைப்படலாம். �
ஆக்ஸிஜனேற்ற அழுத்தம் என்பது ஃப்ரீ ரேடிக்கல்கள் அல்லது நிலையற்ற மூலக்கூறுகளால் ஏற்படும் செல் சேதம் என விவரிக்கப்படுகிறது, இது இறுதியில் ஆரோக்கியமான செயல்பாட்டை பாதிக்கும். பாக்டீரியா மற்றும் வைரஸ்களை நடுநிலையாக்க மனித உடல் ஃப்ரீ ரேடிக்கல்களை உருவாக்குகிறது, இருப்பினும், ஆக்ஸிஜன், மாசுபாடு மற்றும் கதிர்வீச்சு போன்ற வெளிப்புற காரணிகளும் பெரும்பாலும் ஃப்ரீ ரேடிக்கல்களை உருவாக்கலாம். ஆக்ஸிஜனேற்ற அழுத்தம் பல உடல்நலப் பிரச்சினைகளுடன் தொடர்புடையது.
ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் பிற அழுத்தங்கள் மனித உடலின் ஆக்ஸிஜனேற்ற பதிலைக் கட்டுப்படுத்த உதவும் உள் பாதுகாப்பு வழிமுறைகளை இயக்குகின்றன. Nrf2 என்பது ஒரு புரதமாகும், இது ஆக்ஸிஜனேற்ற அழுத்தத்தின் அளவை உணர்ந்து செல்கள் உள் மற்றும் வெளிப்புற காரணிகளிலிருந்து தங்களைப் பாதுகாத்துக் கொள்ள உதவுகிறது. ஆக்ஸிஜனேற்ற நொதிகள் மற்றும் அழுத்த-பதில் மரபணுக்களின் உற்பத்தியில் ஈடுபட்டுள்ள மரபணுக்களை ஒழுங்குபடுத்துவதற்கு Nrf2 உதவுவதாகவும் நிரூபிக்கப்பட்டுள்ளது. என்பதை விளக்குவதே கீழே உள்ள கட்டுரையின் நோக்கம் Nrf2 இன் விளைவுகள் புற்றுநோயில்.
சுருக்கம்
Keap1-Nrf2 பாதை என்பது ஆக்ஸிஜனேற்ற மற்றும் எலக்ட்ரோஃபிலிக் அழுத்தத்திற்கான சைட்டோபுரோடெக்டிவ் பதில்களின் முக்கிய சீராக்கி ஆகும். டிரான்ஸ்கிரிப்ஷன் காரணி Nrf2 மூலம் தூண்டப்பட்ட செல் சிக்னலிங் பாதைகள் சாதாரண மற்றும் முன்கூட்டிய திசுக்களில் புற்றுநோய் ஆரம்பம் மற்றும் முன்னேற்றத்தைத் தடுக்கின்றன, முழு வீரியம் மிக்க உயிரணுக்களில் Nrf2 செயல்பாடு புற்றுநோய் வேதியியல் தன்மையை அதிகரிப்பதன் மூலம் மற்றும் கட்டி உயிரணு வளர்ச்சியை அதிகரிப்பதன் மூலம் வளர்ச்சி நன்மையை வழங்குகிறது. இந்த வரைகலை மதிப்பாய்வில், Keap1-Nrf2 பாதை மற்றும் புற்றுநோய் உயிரணுக்களில் அதன் ஒழுங்குபடுத்தல் பற்றிய கண்ணோட்டத்தை வழங்குகிறோம். புற்றுநோய் உயிரணுக்களில் உள்ள Nrf2 செயல்பாட்டின் விளைவுகளையும், புற்றுநோய் மரபணு சிகிச்சையில் இதை எவ்வாறு பயன்படுத்தலாம் என்பதையும் சுருக்கமாகச் சுருக்கமாகக் கூறுகிறோம்.
முக்கிய வார்த்தைகள்:Nrf2, Keap1, புற்றுநோய், ஆக்ஸிஜனேற்ற மறுமொழி உறுப்பு, மரபணு சிகிச்சை
அறிமுகம்
Keap1-Nrf2 பாதையானது எதிர்வினை ஆக்ஸிஜன் இனங்கள் (ROS) மற்றும் எலக்ட்ரோஃபைல்கள் [1] ஆகியவற்றால் ஏற்படும் உட்புற மற்றும் வெளிப்புற அழுத்தங்களுக்கு சைட்டோபுரோடெக்டிவ் பதில்களின் முக்கிய சீராக்கி ஆகும். பாதையில் உள்ள முக்கிய சமிக்ஞை புரதங்கள் டிரான்ஸ்கிரிப்ஷன் காரணி Nrf2 (அணு காரணி எரித்ராய்டு 2-தொடர்புடைய காரணி 2) ஆகும், இது சிறிய மாஃப் புரதங்களுடன் இலக்கு மரபணுக்களின் ஒழுங்குமுறை பகுதிகளில் உள்ள ஆக்ஸிஜனேற்ற மறுமொழி உறுப்பு (ARE) உடன் பிணைக்கிறது, மற்றும் Keap1 (Kelch ECH அசோசியேட்டிங் புரோட்டீன் 1), ஒரு அடக்குமுறை புரதம் Nrf2 உடன் பிணைக்கிறது மற்றும் எபிக்விடின் புரோட்டீசோம் பாதையால் அதன் சிதைவை ஊக்குவிக்கிறது (படம் 1). கீப்1 மிகவும் சிஸ்டைன் நிறைந்த புரதமாகும், மவுஸ் கீப்1 மொத்தம் 25 மற்றும் மனித 27 சிஸ்டைன் எச்சங்களைக் கொண்டுள்ளது, இவற்றில் பெரும்பாலானவை வெவ்வேறு ஆக்ஸிஜனேற்றங்கள் மற்றும் எலக்ட்ரோஃபைல்கள் மூலம் விட்ரோவில் மாற்றியமைக்கப்படலாம் [2]. இந்த மூன்று எச்சங்கள், C151, C273 மற்றும் C288, Keap1 இன் இணக்கத்தை மாற்றுவதன் மூலம் Nrf2 இன் அணுக்கரு இடமாற்றம் மற்றும் அடுத்தடுத்த இலக்கு மரபணு வெளிப்பாடு [3] (படம். 1) ஆகியவற்றின் மூலம் ஒரு செயல்பாட்டுப் பாத்திரத்தை வகிக்கின்றன. Keap1 இல் உள்ள சிஸ்டைன் மாற்றங்கள் Nrf2 செயல்பாட்டிற்கு வழிவகுக்கும் சரியான வழிமுறை தெரியவில்லை, ஆனால் இரண்டு நடைமுறையில் இருக்கும் ஆனால் ஒன்றுக்கொன்று பிரத்தியேகமாக இல்லாத மாதிரிகள் (1) கீல் மற்றும் லாட்ச் மாதிரி ஆகும், இதில் Keap1 இன் IVR இல் உள்ள தியோல் எச்சங்களில் Keap1 மாற்றங்கள் செய்யப்படுகின்றன. Nrf2 உடனான தொடர்புகளை சீர்குலைத்து, Nrf2 க்குள் உள்ள லைசின் எச்சங்களை இனி பாலியூபிக்விட்டினிலேட்டாக மாற்ற முடியாது மற்றும் (2) தியோல் மாற்றமானது Keap3 [1] இலிருந்து Cul3 இன் விலகலை ஏற்படுத்தும் மாதிரி. இரண்டு மாதிரிகளிலும், தூண்டி-மாற்றியமைக்கப்பட்ட மற்றும் Nrf2-பிணைக்கப்பட்ட Keap1 செயலிழக்கப்பட்டது, அதன் விளைவாக, புதிதாக ஒருங்கிணைக்கப்பட்ட Nrf2 புரதங்கள் Keap1 ஐத் தவிர்த்து, கருவுக்குள் இடமாற்றம் செய்து, ARE உடன் பிணைக்கப்பட்டு NAD(P)H போன்ற Nrf2 இலக்கு மரபணுக்களின் வெளிப்பாட்டை இயக்குகின்றன. குயினோன் ஆக்சிடோரேடக்டேஸ் 1 (NQO1), ஹீம் ஆக்சிஜனேஸ் 1 (HMOX1), குளுட்டமேட்-சிஸ்டைன் லிகேஸ் (GCL) மற்றும் குளுதாதயோன் S பரிமாற்றங்கள் (GSTகள்) (படம் 2). Nrf1 இலக்கு மரபணு தூண்டலின் விளைவாக Keap2 thiols மாற்றங்களைத் தவிர, p21 மற்றும் p62 போன்ற புரதங்கள் Nrf2 அல்லது Keap1 உடன் பிணைக்கப்படலாம், இதனால் Nrf2 மற்றும் Keap1 [1], [3] (படம் 3) இடையேயான தொடர்புகளை சீர்குலைக்கும்.
படம் 1. Nrf2 மற்றும் Keap1 மற்றும் சிஸ்டைன் குறியீடு ஆகியவற்றின் கட்டமைப்புகள். (A) Nrf2 589 அமினோ அமிலங்களைக் கொண்டுள்ளது மற்றும் ஆறு பரிணாம ரீதியாக மிகவும் பாதுகாக்கப்பட்ட களங்களைக் கொண்டுள்ளது, Neh1-6. Neh1 ஒரு bZip மையக்கருத்தைக் கொண்டுள்ளது, இது ஒரு அடிப்படைப் பகுதி - leucine zipper (L-Zip) அமைப்பு, இதில் அடிப்படைப் பகுதி DNA அங்கீகாரத்திற்குப் பொறுப்பாகும் மற்றும் L-Zip சிறிய மாஃப் புரதங்களுடன் டைமரைசேஷனை மத்தியஸ்தம் செய்கிறது. நியூக்ளியஸில் Nrf6 இன் சிதைவை மத்தியஸ்தம் செய்ய Neh2 ஒரு டிக்ரோனாக செயல்படுகிறது. Neh4 மற்றும் 5 பரிமாற்ற டொமைன்கள். Neh2 இல் ETGE மற்றும் DLG மையக்கருத்துகள் உள்ளன, அவை Keap1 உடனான தொடர்புக்குத் தேவைப்படுகின்றன, மேலும் லைசின் எச்சங்களின் (7 K) ஹைட்ரோஃபிலிக் பகுதி, இது Keap1-சார்ந்த பாலியூபிகுடினேஷன் மற்றும் Nrf2 இன் சீரழிவுக்கு இன்றியமையாதது. (B) Keap1 624 அமினோ அமில எச்சங்களைக் கொண்டுள்ளது மற்றும் ஐந்து களங்களைக் கொண்டுள்ளது. BTB டொமைன் மற்றும் கெல்ச் டொமைன் ஆகிய இரண்டு புரோட்டீன்-புரத தொடர்பு மையக்கருத்துகள் இடைப்பட்ட பகுதியால் (IVR) பிரிக்கப்படுகின்றன. BTB டொமைன் மற்றும் IVR இன் N-டெர்மினல் பகுதியுடன் சேர்ந்து Keap1 இன் ஹோமோடைமரைசேஷன் மற்றும் Cullin3 (Cul3) உடன் பிணைக்கிறது. Kelch டொமைன் மற்றும் C-டெர்மினல் பகுதி ஆகியவை Neh2 உடனான தொடர்புக்கு மத்தியஸ்தம் செய்கின்றன. (C) Nrf2 அதன் Neh1 ETGE மற்றும் DLG மையக்கருத்துகள் மூலம் Keap2 இன் இரண்டு மூலக்கூறுகளுடன் தொடர்பு கொள்கிறது. ETGE மற்றும் DLG இரண்டும் Keap1 Kelch மையக்கருத்தின் கீழ் மேற்பரப்பில் உள்ள ஒத்த தளங்களுடன் பிணைக்கப்படுகின்றன. (D) Keap1 மனித புரதத்தில் 27 சிஸ்டைன்களுடன், சிஸ்டைன் எச்சங்களில் நிறைந்துள்ளது. இந்த சிஸ்டைன்களில் சில அடிப்படை எச்சங்களுக்கு அருகில் அமைந்துள்ளன, எனவே அவை எலக்ட்ரோஃபைல்கள் மற்றும் ஆக்ஸிஜனேற்றங்களின் சிறந்த இலக்குகளாகும். எலக்ட்ரோஃபைல்களால் சிஸ்டைன் எச்சங்களை மாற்றியமைக்கும் முறை சிஸ்டைன் குறியீடு என அழைக்கப்படுகிறது. சிஸ்டைன் குறியீடு கருதுகோள் கட்டமைப்பு ரீதியாக வேறுபட்ட Nrf2 செயல்படுத்தும் முகவர்கள் வெவ்வேறு Keap1 சிஸ்டைன்களை பாதிக்கிறது என்று முன்மொழிகிறது. Nrf1 DLG மற்றும் Keap2 Kelch டொமைன்களுக்கிடையேயான தொடர்புகளை சீர்குலைக்கும் கீப்1 இல் சிஸ்டைன் மாற்றங்கள் இணக்கமான மாற்றங்களுக்கு இட்டுச் செல்கின்றன, இதனால் Nrf2 இன் பாலியூபிகுட்டினேஷனைத் தடுக்கிறது. Cys151, Cys273 மற்றும் Cys288 ஆகியவற்றின் செயல்பாட்டு முக்கியத்துவம் காட்டப்பட்டுள்ளது, ஏனெனில் Nrf273 ஐ அடக்குவதற்கு Cys288 மற்றும் Cys2 மற்றும் தூண்டிகள் [151], [2] மூலம் Nrf1 ஐ செயல்படுத்துவதற்கு Cys3 தேவைப்படுகிறது.
படம் 2. Nrf2-Keap1 சமிக்ஞை பாதை. (A மற்றும் B) அடிப்படை நிலைகளில், இரண்டு Keap1 மூலக்கூறுகள் Nrf2 உடன் பிணைக்கப்படுகின்றன மற்றும் Nrf2 ஆகியவை Cul3-அடிப்படையிலான E3 லிகேஸ் வளாகத்தால் பல்வகைப்படுத்தப்படுகின்றன. இந்த பாலியூபிக்டிலேஷன் புரோட்டீசோம் மூலம் விரைவான Nrf2 சிதைவை ஏற்படுத்துகிறது. Nrf2 இன் ஒரு சிறிய பகுதியானது தடுப்பு வளாகத்திலிருந்து தப்பித்து, அடிப்படை ARE-சார்ந்த மரபணு வெளிப்பாட்டிற்கு மத்தியஸ்தம் செய்ய கருவில் குவிந்து, அதன் மூலம் செல்லுலார் ஹோமியோஸ்டாசிஸை பராமரிக்கிறது. (சி) மன அழுத்த சூழ்நிலையில், தூண்டிகள் கீப்1 சிஸ்டைன்களை மாற்றியமைக்கின்றன, இது தடுப்பு வளாகத்தின் விலகல் மூலம் Nrf2 எங்கும் பரவுவதைத் தடுக்கிறது. (D) கீல் மற்றும் தாழ்ப்பாளை மாதிரியின் படி, குறிப்பிட்ட Keap1 சிஸ்டைன் எச்சங்களை மாற்றியமைப்பது Keap1 இல் இணக்கமான மாற்றங்களுக்கு வழிவகுக்கிறது, இதன் விளைவாக Keap2 இலிருந்து Nrf1 DLG மையக்கருத்தை பிரிக்கிறது. Nrf2 இன் எங்கும் பரவுவது சீர்குலைந்தது ஆனால் ETGE மையக்கருத்துடன் பிணைப்பு உள்ளது. (E) Keap1-Cul3 விலகல் மாதிரியில், எலக்ட்ரோஃபைல்களுக்கு பதிலளிக்கும் விதமாக Keap1 மற்றும் Cul3 பிணைப்பு சீர்குலைந்து, எங்கும் பரவும் அமைப்பிலிருந்து Nrf2 தப்பிக்க வழிவகுக்கிறது. பரிந்துரைக்கப்பட்ட இரண்டு மாதிரிகளிலும், தூண்டி-மாற்றியமைக்கப்பட்ட மற்றும் Nrf2-பிணைக்கப்பட்ட Keap1 செயலிழக்கப்பட்டது, அதன் விளைவாக, புதிதாக ஒருங்கிணைக்கப்பட்ட Nrf2 புரதங்கள் Keap1 ஐத் தவிர்த்து, அணுக்கருவிற்குள் இடமாற்றம் செய்து, ஆக்ஸிஜனேற்ற மறுமொழி உறுப்புடன் (ARE) பிணைக்கப்பட்டு Nrf2 இன் இலக்கு வெளிப்பாட்டை இயக்குகின்றன. NQO1, HMOX1, GCL மற்றும் GSTகள் [1], [3] போன்ற மரபணுக்கள்.
படம் 3. புற்று நோயில் Nrf2 என்ற அணுக்கரு குவிப்புக்கான வழிமுறைகள். (A) Nrf2 அல்லது Keap1 இல் உள்ள சோமாடிக் பிறழ்வுகள் இந்த இரண்டு புரதங்களின் தொடர்புகளை சீர்குலைக்கின்றன. Nrf2 இல், பிறழ்வுகள் ETGE மற்றும் DLG மையக்கருத்துகளைப் பாதிக்கின்றன, ஆனால் Keap1 இல் பிறழ்வுகள் மிகவும் சமமாக விநியோகிக்கப்படுகின்றன. மேலும், KrasG12D[5] போன்ற ஆன்கோஜீன் செயல்படுத்தல் அல்லது PTEN [11] போன்ற கட்டி அடக்கிகளின் சீர்குலைவு Nrf2 இன் டிரான்ஸ்கிரிப்ஷனல் தூண்டல் மற்றும் அணுக்கரு Nrf2 இன் அதிகரிப்புக்கு வழிவகுக்கும். (B) நுரையீரல் மற்றும் புரோஸ்டேட் புற்றுநோயில் உள்ள கீப்1 ஊக்குவிப்பாளரின் ஹைபர்மெதிலேஷன் கீப்1 எம்ஆர்என்ஏ வெளிப்பாட்டைக் குறைக்க வழிவகுக்கிறது, இது என்ஆர்எஃப்2 [6], [7] அணுக் குவிப்பை அதிகரிக்கிறது. (சி) குடும்ப பாப்பில்லரி சிறுநீரக புற்றுநோயில், ஃபுமரேட் ஹைட்ராடேஸ் என்சைம் செயல்பாட்டின் இழப்பு ஃபுமரேட்டின் திரட்சிக்கு வழிவகுக்கிறது மற்றும் மேலும் கீப்1 சிஸ்டைன் எச்சங்கள் (2SC) உறிஞ்சப்படுவதற்கு வழிவகுக்கிறது. இந்த மொழிபெயர்ப்புக்குப் பிந்தைய மாற்றம் Keap1-Nrf2 தொடர்பு மற்றும் Nrf2 [8], [9] இன் அணுக் குவிப்புக்கு இடையூறு விளைவிக்கும். (D) p62 மற்றும் p21 போன்ற சீர்குலைக்கும் புரதங்களின் திரட்சி Nrf2-Keap1 பிணைப்பைத் தொந்தரவு செய்யலாம் மற்றும் அணுக்கரு Nrf2 இல் அதிகரிப்புக்கு வழிவகுக்கும். p62 ஆனது Keap1 உடன் பிணைக்கிறது, Nrf2 க்கான பைண்டிங் பாக்கெட்டை ஒன்றுடன் ஒன்று இணைக்கிறது மற்றும் p21 நேரடியாக Nrf2 இன் DLG மற்றும் ETGE மையக்கருத்துக்களுடன் தொடர்பு கொள்கிறது, இதன் மூலம் Keap1 உடன் போட்டியிடுகிறது [10].
புற்றுநோயில் Nrf2 ஐ செயல்படுத்துதல் மற்றும் ஒழுங்குபடுத்தும் வழிமுறைகள்
Nrf2 ஆக்டிவேஷன் மூலம் வழங்கப்படும் சைட்டோபுரோடெக்ஷன் சாதாரண மற்றும் முன்கூட்டிய திசுக்களில் புற்றுநோய் வேதியியல் தடுப்புக்கு முக்கியமானது என்றாலும், முழு வீரியம் மிக்க உயிரணுக்களில் Nrf2 செயல்பாடு புற்றுநோய் வேதியியல் தன்மையை அதிகரிப்பதன் மூலமும் கட்டி உயிரணு வளர்ச்சியை அதிகரிப்பதன் மூலமும் வளர்ச்சி நன்மையை வழங்குகிறது [4]. பல்வேறு புற்றுநோய்களில் Nrf2 சிக்னலிங் பாதை அமைப்புரீதியாக செயல்படுத்தப்படும் பல வழிமுறைகள் விவரிக்கப்பட்டுள்ளன: (1) Keap1 இல் உள்ள உடலியல் பிறழ்வுகள் அல்லது Nrf1 இன் Keap2 பிணைப்பு டொமைன் அவற்றின் தொடர்புகளை சீர்குலைக்கிறது; (2) Keap1 வெளிப்பாட்டின் எபிஜெனெடிக் சைலன்சிங் Nrf2 இன் குறைபாடுள்ள அடக்குமுறைக்கு வழிவகுக்கிறது; (3) கீப்62-என்ஆர்எஃப்1 வளாகத்தின் விலகலுக்கு வழிவகுக்கும் p2 போன்ற சீர்குலைக்கும் புரதங்களின் குவிப்பு; (4) ஆன்கோஜெனிக் கே-ராஸ், பி-ராஃப் மற்றும் சி-மைக் மூலம் Nrf2 இன் டிரான்ஸ்கிரிப்ஷனல் தூண்டல்; மற்றும் (5) ஃபுமரேட் ஹைட்ரேடேஸ் என்சைம் செயல்பாடு [1], [3], [4], [5], [6], [7], [8], [9], [10], [3], [2] 2], [4], [2] (படம் 11). அமைப்புரீதியாக ஏராளமாக உள்ள Nrf4 புரதம், மருந்து வளர்சிதை மாற்றத்தில் ஈடுபட்டுள்ள மரபணுக்களின் வெளிப்பாட்டை அதிகரிக்கச் செய்கிறது, இதனால் வேதிச்சிகிச்சை மருந்துகள் மற்றும் கதிரியக்க சிகிச்சைக்கான எதிர்ப்பை அதிகரிக்கிறது. கூடுதலாக, உயர் NrfXNUMX புரத அளவு புற்றுநோயின் மோசமான முன்கணிப்புடன் தொடர்புடையது [XNUMX]. அதிகப்படியான NrfXNUMX, குளுக்கோஸ் மற்றும் குளுட்டமைனை அனபோலிக் பாதைகளை நோக்கி செலுத்துவதன் மூலம் செல் பெருக்கத்தையும் பாதிக்கிறது, இது பியூரின் தொகுப்பை அதிகரிக்கிறது மற்றும் செல் பெருக்கத்தை ஊக்குவிக்க பென்டோஸ் பாஸ்பேட் பாதையை பாதிக்கிறது [XNUMX] (படம். XNUMX).
படம் 4. டூமோரிஜெனெசிஸில் Nrf2 இன் இரட்டைப் பங்கு. உடலியல் நிலைமைகளின் கீழ், செல்லுலார் ஹோமியோஸ்டாசிஸை பராமரிக்க குறைந்த அளவிலான அணு Nrf2 போதுமானது. Nrf2 புற்றுநோய்கள், ROS மற்றும் பிற டிஎன்ஏ-சேதமடைந்த முகவர்களை நீக்குவதன் மூலம் கட்டி தொடங்குதல் மற்றும் புற்றுநோய் மெட்டாஸ்டாசிஸைத் தடுக்கிறது. டூமோரிஜெனெசிஸின் போது, டிஎன்ஏ சேதத்தை குவிப்பது Nrf2 இன் கான்ஸ்டிட்யூட்டிவ் ஹைபராக்டிவிட்டிக்கு வழிவகுக்கிறது, இது தன்னாட்சி வீரியம் மிக்க செல்கள் அதிக அளவு எண்டோஜெனஸ் ROS ஐத் தாங்கி, அப்போப்டொசிஸைத் தவிர்க்க உதவுகிறது. தொடர்ந்து உயர்த்தப்பட்ட அணுக்கரு Nrf2 அளவுகள் வளர்சிதை மாற்ற மரபணுக்களை செயல்படுத்துகின்றன, மேலும் சைட்டோபுரோடெக்டிவ் மரபணுக்களுடன் வளர்சிதை மாற்ற மறுசீரமைப்பு மற்றும் மேம்பட்ட செல் பெருக்கத்திற்கு பங்களிக்கின்றன. அதிக Nrf2 அளவுகளைக் கொண்ட புற்றுநோய்கள் மோசமான முன்கணிப்புடன் தொடர்புடையவை, ஏனெனில் ரேடியோ மற்றும் வேதியியல் தன்மை மற்றும் ஆக்கிரமிப்பு புற்றுநோய் செல் பெருக்கம். எனவே, டூமோரிஜெனிசிஸின் ஆரம்ப கட்டங்களில் Nrf2 பாதையின் செயல்பாடு பாதுகாப்பாக உள்ளது, ஆனால் பிந்தைய நிலைகளில் தீங்கு விளைவிக்கும். எனவே, புற்றுநோயைத் தடுப்பதற்கு, Nrf2 செயல்பாட்டை மேம்படுத்துவது ஒரு முக்கியமான அணுகுமுறையாக உள்ளது, அதேசமயம் புற்றுநோய் சிகிச்சைக்கு, Nrf2 தடுப்பு விரும்பத்தக்கது [4], [11].
உயர் Nrf2 செயல்பாடு பொதுவாக புற்றுநோய் உயிரணுக்களில் பாதகமான விளைவுகளுடன் நிகழ்கிறது என்பதால், Nrf2 ஐத் தடுக்க சிகிச்சைகள் தேவைப்படுகின்றன. துரதிர்ஷ்டவசமாக, வேறு சில bZip குடும்ப உறுப்பினர்களுடனான கட்டமைப்பு ஒற்றுமை காரணமாக, குறிப்பிட்ட Nrf2 தடுப்பான்களின் வளர்ச்சி ஒரு சவாலான பணியாகும், மேலும் Nrf2 தடுப்பு பற்றிய சில ஆய்வுகள் மட்டுமே இன்றுவரை வெளியிடப்பட்டுள்ளன. இயற்கை தயாரிப்புகளை திரையிடுவதன் மூலம், ரென் மற்றும் பலர். [12] சிஸ்ப்ளேட்டின் கீமோதெரபியூடிக் செயல்திறனை மேம்படுத்தும் Nrf2 இன்ஹிபிட்டராக ஆன்டினியோபிளாஸ்டிக் கலவை புருசடோலை அடையாளம் கண்டுள்ளது. கூடுதலாக, PI3K தடுப்பான்கள் [11], [13] மற்றும் Nrf2 siRNA [14] ஆகியவை புற்றுநோய் உயிரணுக்களில் Nrf2 ஐத் தடுக்கப் பயன்படுத்தப்படுகின்றன. சமீபத்தில், உயர் Nrf2 அளவுகள் கொண்ட புற்றுநோய் செல்களை குறிவைக்க, புற்றுநோய் தற்கொலை மரபணு சிகிச்சை எனப்படும் மாற்று அணுகுமுறையைப் பயன்படுத்தினோம். தைமிடின் கைனேஸ் (TK) கொண்ட Nrf2-உந்துதல் லென்டிவைரல் திசையன்கள் [15] உயர் ARE செயல்பாடு கொண்ட புற்றுநோய் உயிரணுக்களுக்கு மாற்றப்படுகின்றன மற்றும் செல்கள் ஒரு சார்பு மருந்து, கான்சிக்ளோவிர் (GCV) மூலம் சிகிச்சையளிக்கப்படுகின்றன. GCV ஆனது GCV-மோனோபாஸ்பேட்டாக வளர்சிதைமாற்றம் செய்யப்படுகிறது, இது செல்லுலார் கைனேஸ்களால் மேலும் பாஸ்போரிலேட் செய்யப்பட்டு நச்சு ட்ரைபாஸ்பேட் வடிவத்தில் [16] (படம் 5). இது TK ஐக் கொண்டிருக்கும் கட்டி செல்களை மட்டுமல்ல, பார்வையாளர் விளைவின் காரணமாக அண்டை செல்களையும் திறம்பட கொல்ல வழிவகுக்கிறது [17]. ARE-ஒழுங்குபடுத்தப்பட்ட TK/GCV மரபணு சிகிச்சையானது புற்றுநோய் வேதியியல் சிகிச்சை முகவரான டாக்ஸோரூபிகின் சிகிச்சையுடன் [16] இணைப்பதன் மூலம் மேலும் மேம்படுத்தப்படலாம், இந்த அணுகுமுறை பாரம்பரிய சிகிச்சைகளுடன் இணைந்து பயனுள்ளதாக இருக்கும் என்ற கருத்தை ஆதரிக்கிறது.
படம் 5. தற்கொலை மரபணு சிகிச்சை. புற்றுநோய் உயிரணுக்களில் உள்ள Nrf2 அணுக்கரு குவிப்பை புற்றுநோய் தற்கொலை மரபணு சிகிச்சைக்காக Nrf2-உந்துதல் வைரஸ் திசையன் பயன்படுத்துவதன் மூலம் பயன்படுத்திக் கொள்ளலாம் [16]. இந்த அணுகுமுறையில், நான்கு AREகளுடன் குறைந்தபட்ச SV40 ஊக்குவிப்பாளரின் கீழ் தைமிடின் கைனேஸை (TK) வெளிப்படுத்தும் லென்டிவைரல் வெக்டார் (LV) நுரையீரல் அடினோகார்சினோமா செல்களுக்கு கடத்தப்படுகிறது. உயர் அணுக்கரு Nrf2 நிலைகள் Nrf2 பிணைப்பு மூலம் TK இன் வலுவான வெளிப்பாட்டிற்கு வழிவகுக்கும். செல்கள் பின்னர் TK ஆல் பாஸ்போரிலேட்டட் செய்யப்பட்ட கான்சிக்ளோவிர் (GCV) என்ற சார்பு மருந்துடன் சிகிச்சையளிக்கப்படுகின்றன. டிரைஃபோஸ்ஃபோரிலேட்டட் ஜிசிவி டிஎன்ஏ தொகுப்பை சீர்குலைக்கிறது மற்றும் டிகேவைக் கொண்ட கட்டி செல்களை மட்டுமல்ல, பார்வையாளர் விளைவின் காரணமாக அண்டை செல்களையும் திறம்பட கொல்ல வழிவகுக்கிறது.
Nrf2 என்பது மனித உடலில் சக்திவாய்ந்த ஆக்ஸிஜனேற்ற உற்பத்தியைத் தூண்டும் ஒரு முதன்மை சீராக்கி ஆகும், இது ஆக்ஸிஜனேற்ற அழுத்தத்தை அகற்ற உதவுகிறது. சூப்பர் ஆக்சைடு டிஸ்முடேஸ், அல்லது SOD, குளுதாதயோன் மற்றும் கேடலேஸ் போன்ற பல்வேறு ஆக்ஸிஜனேற்ற நொதிகளும் Nrf2 பாதை வழியாக செயல்படுத்தப்படுகின்றன. மேலும், மஞ்சள், அஸ்வகந்தா, பக்கோபா, கிரீன் டீ மற்றும் பால் திஸ்டில் போன்ற சில தாவர வேதிப்பொருட்கள் Nrf2 ஐ செயல்படுத்துகின்றன. என்று ஆய்வுகள் கண்டறிந்துள்ளன Nrf2 செயல்படுத்தல் இயற்கையாகவே செல்லுலார் பாதுகாப்பை மேம்படுத்தி மனித உடலுக்கு சமநிலையை மீட்டெடுக்க முடியும்.
டாக்டர். அலெக்ஸ் ஜிமினெஸ் DC, CCST இன்சைட்
புற்றுநோய், இறப்பு, முதுமை, மூளை மற்றும் நடத்தை, இதய நோய் மற்றும் பலவற்றில் சல்போராபேன் மற்றும் அதன் விளைவுகள்
ஐசோதியோசயனேட்டுகள் உங்கள் உணவில் நீங்கள் பெறக்கூடிய மிக முக்கியமான தாவர கலவைகள் ஆகும். இந்த வீடியோவில் நான் அவர்களுக்காக இதுவரை செய்யப்படாத மிக விரிவான வழக்கை உருவாக்குகிறேன். குறுகிய கவனம்? கீழே உள்ள நேரப் புள்ளிகளில் ஒன்றைக் கிளிக் செய்வதன் மூலம் உங்களுக்குப் பிடித்த தலைப்புக்குச் செல்லவும். முழு காலவரிசை கீழே.
முக்கிய பிரிவுகள்:
00:01:14 - புற்றுநோய் மற்றும் இறப்பு
00:19:04 - முதுமை
00:26:30 - மூளை மற்றும் நடத்தை
00:38:06 - இறுதி மறுபரிசீலனை
00:40:27 - டோஸ்
முழு காலவரிசை:
00:00:34 – வீடியோவின் முக்கிய மையமான சல்போராபேன் அறிமுகம்.
00:01:14 - க்ரூசிஃபெரஸ் காய்கறி நுகர்வு மற்றும் அனைத்து காரணங்களின் இறப்பு குறைப்பு.
00:02:12 - புரோஸ்டேட் புற்றுநோய் ஆபத்து.
00:02:23 - சிறுநீர்ப்பை புற்றுநோய் ஆபத்து.
00:02:34 - புகைப்பிடிப்பவர்களுக்கு நுரையீரல் புற்றுநோய் ஆபத்து.
00:02:48 - மார்பக புற்றுநோய் ஆபத்து.
00:03:13 - அனுமானம்: உங்களுக்கு ஏற்கனவே புற்றுநோய் இருந்தால் என்ன செய்வது? (இடையிடல்)
00:03:35 – புற்றுநோய் மற்றும் இறப்பு தொடர்பான தரவுகளை இயக்கும் நம்பத்தகுந்த வழிமுறை.
00:04:38 - சல்போராபேன் மற்றும் புற்றுநோய்.
00:05:32 - எலிகளில் சிறுநீர்ப்பை கட்டி வளர்ச்சியில் ப்ரோக்கோலி முளை சாறு வலுவான விளைவைக் காட்டும் விலங்கு சான்றுகள்.
00:06:06 - புரோஸ்டேட் புற்றுநோயாளிகளில் சல்ஃபோராபேன் நேரடியாக நிரப்புவதன் விளைவு.
00:07:09 - உண்மையான மார்பக திசுக்களில் ஐசோதியோசயனேட் வளர்சிதை மாற்றங்களின் உயிர் குவிப்பு.
00:08:32 - மார்பக புற்றுநோய் ஸ்டெம் செல்களைத் தடுப்பது.
00:08:53 - வரலாற்றுப் பாடம்: பண்டைய ரோமில் கூட பிராசிகாக்கள் ஆரோக்கிய பண்புகளைக் கொண்டிருப்பதாக நிறுவப்பட்டது.
00:09:16 – சல்போராபேன் புற்றுநோய் வெளியேற்றத்தை மேம்படுத்தும் திறன் (பென்சீன், அக்ரோலின்).
00:09:51 - ஆன்டிஆக்ஸிடன்ட் மறுமொழி கூறுகள் வழியாக மரபணு மாற்றமாக NRF2.
00:10:10 – NRF2 ஆக்டிவேஷன் குளுதாதயோன்-எஸ்-கான்ஜுகேட்ஸ் வழியாக புற்றுநோயை வெளியேற்றுவதை எவ்வாறு மேம்படுத்துகிறது.
00:10:34 - பிரஸ்ஸல்ஸ் முளைகள் குளுதாதயோன்-எஸ்-டிரான்ஸ்ஃபெரேஸை அதிகரிக்கின்றன மற்றும் டிஎன்ஏ பாதிப்பைக் குறைக்கின்றன.
00:11:20 - ப்ரோக்கோலி முளை பானம் பென்சீன் வெளியேற்றத்தை 61% அதிகரிக்கிறது.
00:13:31 - ப்ரோக்கோலி ஸ்ப்ரூட் ஹோமோஜெனேட் மேல் சுவாசப்பாதையில் ஆக்ஸிஜனேற்ற நொதிகளை அதிகரிக்கிறது.
00:15:45 - சிலுவை காய்கறி நுகர்வு மற்றும் இதய நோய் இறப்பு.
00:16:55 - ப்ரோக்கோலி ஸ்ப்ரூட் பவுடர் இரத்த கொழுப்பு மற்றும் ஒட்டுமொத்த இதய நோய் அபாயத்தை மேம்படுத்துகிறது வகை 2 நீரிழிவு நோயாளிகள்.
00:19:04 - வயதான பிரிவின் ஆரம்பம்.
00:19:21 - சல்போராபேன்-செறிவூட்டப்பட்ட உணவு வண்டுகளின் ஆயுட்காலம் 15 முதல் 30% வரை அதிகரிக்கிறது (சில நிபந்தனைகளில்).
00:20:34 - நீண்ட ஆயுளுக்கு குறைந்த வீக்கத்தின் முக்கியத்துவம்.
00:22:05 - குரூசிஃபெரஸ் காய்கறிகள் மற்றும் ப்ரோக்கோலி ஸ்ப்ரூட் பவுடர் மனிதர்களில் பல்வேறு வகையான அழற்சி குறிப்பான்களைக் குறைப்பதாகத் தெரிகிறது.
00:23:40 – மிட்-வீடியோ ரீகேப்: புற்றுநோய், வயதான பிரிவுகள்
00:24:14 - முதுமையில் சல்போராபேன் தகவமைப்பு நோயெதிர்ப்புச் செயல்பாட்டை மேம்படுத்தலாம் என்று சுட்டி ஆய்வுகள் தெரிவிக்கின்றன.
00:25:18 – சல்ஃபோராபேன், வழுக்கை மவுஸ் மாதிரியில் முடி வளர்ச்சியை மேம்படுத்தியது. படம் 00:26:10.
00:28:17 - மனச்சோர்வு விவாதத்தின் தொடக்கம் (நம்பத்தகுந்த வழிமுறை மற்றும் ஆய்வுகள்).
00:31:21 – அழுத்தத்தால் தூண்டப்பட்ட மனச்சோர்வின் 10 வெவ்வேறு மாதிரிகளைப் பயன்படுத்தி மவுஸ் ஆய்வு, ஃப்ளூக்ஸெடின் (ப்ரோசாக்) போலவே சல்ஃபோராபேன் பயனுள்ளதாக இருக்கும்.
00:32:00 – எலிகளில் குளுக்கோராபனின் நேரடியாக உட்கொள்வது சமூக தோல்வி மன அழுத்த மாதிரியிலிருந்து மனச்சோர்வைத் தடுப்பதில் இதேபோல் பயனுள்ளதாக இருக்கும் என்று ஆய்வு காட்டுகிறது.
00:33:01 - நியூரோடிஜெனரேஷன் பிரிவின் ஆரம்பம்.
00:33:30 – சல்போராபேன் மற்றும் அல்சைமர் நோய்.
00:33:44 – சல்போராபேன் மற்றும் பார்கின்சன் நோய்.
00:33:51 – சல்போராபேன் மற்றும் ஹங்டிங்டன் நோய்.
00:34:13 - சல்ஃபோராபேன் வெப்ப அதிர்ச்சி புரதங்களை அதிகரிக்கிறது.
00:34:43 - அதிர்ச்சிகரமான மூளை காயம் பிரிவின் ஆரம்பம்.
00:35:55 - சல்ஃபோராபேன் மற்றும் நியூரானல் பிளாஸ்டிசிட்டி.
00:36:32 – எலிகளில் டைப் II நீரிழிவு மாதிரியில் சல்போராபேன் கற்றலை மேம்படுத்துகிறது.
00:37:19 - சல்போராபேன் மற்றும் டுச்சேன் தசைநார் சிதைவு.
00:37:44 - தசை செயற்கைக்கோள் செல்களில் Myostatin தடுப்பு (விட்ரோவில்).
00:38:06 – லேட்-வீடியோ மறுபரிசீலனை: இறப்பு மற்றும் புற்றுநோய், டிஎன்ஏ சேதம், ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் வீக்கம், பென்சீன் வெளியேற்றம், இருதய நோய், வகை II நீரிழிவு, மூளையில் விளைவுகள் (மன அழுத்தம், மன இறுக்கம், ஸ்கிசோஃப்ரினியா, நியூரோடிஜெனரேஷன்), NRF2 பாதை.
00:40:27 - ப்ரோக்கோலி முளைகள் அல்லது சல்ஃபோராபேன் அளவைக் கண்டறிவது பற்றிய எண்ணங்கள்.
00:41:01 - வீட்டில் முளைப்பது பற்றிய நிகழ்வுகள்.
00:43:14 - சமையல் வெப்பநிலை மற்றும் சல்ஃபோராபேன் செயல்பாடு.
00:43:45 - குளுகோராபனினில் இருந்து சல்போராபேன் குடல் பாக்டீரியாவை மாற்றுகிறது.
00:44:24 - காய்கறிகளிலிருந்து செயலில் உள்ள மைரோசினேஸுடன் இணைந்தால் சப்ளிமெண்ட்ஸ் சிறப்பாகச் செயல்படும்.
00:44:56 - சமையல் நுட்பங்கள் மற்றும் சிலுவை காய்கறிகள்.
00:46:06 - ஐசோதியோசயனேட்டுகள் கோயிட்ரோஜன்களாக.
அனுமதிகள்
இந்த வேலைக்கு ஃபின்லாந்து அகாடமி, சிக்ரிட் ஜூசிலியஸ் அறக்கட்டளை மற்றும் ஃபின்னிஷ் புற்றுநோய் அமைப்புகள் ஆதரவு அளித்தன.
முடிவில், அணுக்கரு காரணி (எரித்ராய்டு-பெறப்பட்ட 2) போன்ற 2, NFE2L2 அல்லது Nrf2 என்றும் அழைக்கப்படுகிறது, இது ஒரு புரதமாகும், இது ஆக்ஸிஜனேற்ற அழுத்தத்திற்கு எதிராக மனித உடலைப் பாதுகாக்கும் ஆக்ஸிஜனேற்ற உற்பத்தியை அதிகரிக்கிறது. மேலே விவரிக்கப்பட்டுள்ளபடி, புற்றுநோய் உட்பட ஆக்ஸிஜனேற்ற அழுத்தத்தால் ஏற்படும் நோய்களுக்கான சிகிச்சைக்கான Nrf2 பாதையின் தூண்டுதல் ஆய்வுகள் ஆகும். எங்கள் தகவலின் நோக்கம் உடலியக்க மற்றும் முதுகெலும்பு சுகாதார பிரச்சினைகள் மட்டுமே. விஷயத்தைப் பற்றி விவாதிக்க, தயவு செய்து டாக்டர் ஜிமினெஸைக் கேட்கவும் அல்லது எங்களை இங்கே தொடர்பு கொள்ளவும்915-850-0900.
கூடுதல் தலைப்பு விவாதம்: அறுவை சிகிச்சை இல்லாமல் முழங்கால் வலியை நீக்குதல்
முழங்கால் வலி என்பது நன்கு அறியப்பட்ட அறிகுறியாகும், இது பல்வேறு முழங்கால் காயங்கள் மற்றும்/அல்லது நிலைமைகள் காரணமாக ஏற்படலாம்.விளையாட்டு காயங்கள். முழங்கால் மனித உடலில் மிகவும் சிக்கலான மூட்டுகளில் ஒன்றாகும், ஏனெனில் இது நான்கு எலும்புகள், நான்கு தசைநார்கள், பல்வேறு தசைநாண்கள், இரண்டு மெனிசிஸ் மற்றும் குருத்தெலும்பு ஆகியவற்றின் குறுக்குவெட்டுகளால் ஆனது. அமெரிக்கன் அகாடமி ஆஃப் ஃபேமிலி பிசிஷியன்களின் கூற்றுப்படி, முழங்கால் வலிக்கான பொதுவான காரணங்களில் பட்டேலர் சப்லக்சேஷன், பட்டேலர் டெண்டினிடிஸ் அல்லது ஜம்பர்ஸ் முழங்கால் மற்றும் ஓஸ்குட்-ஸ்க்லாட்டர் நோய் ஆகியவை அடங்கும். முழங்கால் வலி 60 வயதுக்கு மேற்பட்டவர்களுக்கு ஏற்பட வாய்ப்புள்ளது என்றாலும், குழந்தைகள் மற்றும் இளம்பருவத்தினருக்கும் முழங்கால் வலி ஏற்படலாம். முழங்கால் வலியை RICE முறைகளைப் பின்பற்றி வீட்டிலேயே சிகிச்சை செய்யலாம், இருப்பினும், கடுமையான முழங்கால் காயங்களுக்கு உடலியக்க சிகிச்சை உட்பட உடனடி மருத்துவ கவனிப்பு தேவைப்படலாம்.
டிஎன்ஏ தோராயமாக 20,000 மரபணுக்களை ஆதரிக்கிறது, ஒவ்வொன்றும் ஆரோக்கியமான வாழ்க்கை முறைக்கு தேவையான புரதம் அல்லது நொதியை உருவாக்குவதற்கான ஒரு திட்டத்தை வைத்திருக்கிறது. இந்த வடிவங்கள் ஒவ்வொன்றும் ஒரு வகையான "ஊக்குவிப்பாளர்" மூலம் தொடர்ந்து ஒழுங்குபடுத்தப்பட வேண்டும், இது ஒவ்வொரு பொருள் மற்றும்/அல்லது இரசாயனங்கள் எவ்வளவு உருவாக்கப்படுகின்றன மற்றும் எந்த நிலைமைகளின் கீழ் அவை உருவாகும் என்பதை சரியாக நிர்வகிக்கிறது.
ஆன்டிஆக்ஸிடன்ட் ரெஸ்பான்ஸ் எலிமென்ட் அல்லது ARE என அழைக்கப்படும் ஒரு குறிப்பிட்ட வகையான சுவிட்ச் போன்ற ஊக்குவிப்பு பகுதிகளுடன் இணைப்பதன் மூலம் Nrf2 காரணிநூற்றுக்கணக்கான தனித்துவமான மரபணுக்களை உருவாக்குவதற்கான வேகத்தை ஆதரிக்கிறது, இது மன அழுத்த சூழ்நிலைகளில் உயிரணுக்களை உயிர்வாழ உதவுகிறது. இந்த மரபணுக்கள் ஆக்ஸிஜனேற்ற நொதிகளின் தேர்வை உருவாக்குகின்றன, அவை ஆக்ஸிஜனேற்றத்தை நடுநிலையாக்குவதன் மூலம் ஒரு பாதுகாப்பு வலையமைப்பை உருவாக்குகின்றன மற்றும் அவற்றின் உற்பத்தியில் விட்டுச்செல்லும் நச்சு துணை தயாரிப்புகளை சுத்தம் செய்வதன் மூலம், அவை ஏற்படுத்திய சேதத்தை மீட்டெடுக்க உதவுகின்றன.
ஆக்ஸிஜனேற்ற அழுத்தம் என்றால் என்ன?
சூப்பர் ஆக்சைடு ரேடிக்கல், அல்லது O2-., மற்றும் ஹைட்ரஜன் பெராக்சைடு, அல்லது H2O2 போன்ற பல ஆக்ஸிஜனேற்றிகள், மனித உடலைத் தாங்கும் பொருட்கள் மற்றும்/அல்லது இரசாயனங்களை எரிக்கும் நடைமுறையின் மூலம் உருவாக்கப்பட்டுள்ளன. மனித உடலில் ஆக்ஸிஜனேற்ற என்சைம்கள் உள்ளன, அவை நாம் உட்கொள்ளும் எதிர்வினை உணவுகள் மற்றும் பானங்களை நடுநிலையாக்குகின்றன மற்றும் நச்சுத்தன்மையாக்குகின்றன. Nrf2 சமநிலையை வைத்திருக்க அவற்றின் உற்பத்தியை மாற்றியமைக்கிறது மற்றும் இந்த நொதிகளின் தேவையை அடிக்கோடிட்டுக் காட்டுகிறது. இந்த சமநிலை வயது உட்பட இரண்டு காரணிகளால் குறுக்கிடப்படலாம்.
நாம் வயதாகும்போது, மனித உடல் குறைவான Nrf2 ஐ உருவாக்குகிறது, மேலும் இந்த நுட்பமான சமநிலை படிப்படியாக ஆக்ஸிஜனேற்ற பக்கத்தை நோக்கி திரும்ப ஆரம்பிக்கும், இது ஆக்ஸிஜனேற்ற அழுத்தம் என குறிப்பிடப்படுகிறது. நோய் ஆக்ஸிஜனேற்றத்தின் அதிகப்படியான உற்பத்தியையும் ஏற்படுத்தக்கூடும். நோய்த்தொற்றுகள், ஒவ்வாமைகள் மற்றும் தன்னுடல் தாக்கக் கோளாறுகள் கூடுதலாக நமது நோயெதிர்ப்பு உயிரணுக்களை O2- போன்ற எதிர்வினை ஆக்ஸிஜனேற்றங்களை உருவாக்கத் தூண்டும். , H2O2, OH மற்றும் HOCl, ஆரோக்கியமான செல்கள் சேதமடைந்து வீக்கத்துடன் பதிலளிக்கின்றன. மாரடைப்பு, பக்கவாதம், புற்றுநோய் மற்றும் அல்சைமர் நோய் போன்ற நரம்பியக்கடத்தல் நிலைமைகள் உட்பட வயதானவுடன் தொடர்புடைய நோய்கள் ஆக்ஸிஜனேற்றத்தின் வளர்ச்சியை அதிகரிக்கின்றன, மன அழுத்தத்தை உருவாக்குகின்றன மற்றும் வீக்கத்தை ஏற்படுத்துகின்றன.
Nrf2 ஆக்டிவேட்டர்கள் என்றால் என்ன?
Nrf2 புரதம், இது என்சைம்கள் மற்றும் மரபணுக்களை ஆதரிக்கும் மற்றும் கட்டுப்படுத்தும் விதத்தின் காரணமாக டிரான்ஸ்கிரிப்ஷன் காரணி என்றும் அழைக்கப்படுகிறது, இது உயிரணுவில் உள்ள உயிர்வேதியியல் எதிர்வினைகளின் வரிசையின் ரகசிய உறுப்பு ஆகும், இது அறிவாற்றல் சமநிலை மற்றும் ஆக்ஸிஜனேற்ற சமநிலையில் ஏற்படும் மாற்றங்களுக்கு வினைபுரிகிறது. இந்த பாதையின் உணர்திறன் கூறுகள் Nrf2 ஐ மாற்றியமைத்து வெளியேற்றுகின்றன, இது டிஎன்ஏவை நோக்கி செல்லின் கருவுக்குள் பரவக்கூடும். Nrf2, உயிரணுவைப் பாதுகாக்க அது ஆதரிக்கும் மரபணுக்கள் மற்றும் என்சைம்களை மாற்றாக இயக்கலாம் அல்லது அணைக்கலாம்.
அதிர்ஷ்டவசமாக, பல நூற்றாண்டுகளுக்கு முன்பு சீன மற்றும் பூர்வீக அமெரிக்க பாரம்பரிய மருந்துகளில் பயன்படுத்தப்பட்ட சில தாவரங்கள் மற்றும் சாறுகளின் நுகர்வு மூலம் Nrf2 ஆக்டிவேட்டர்களான பல்வேறு பொருட்கள் உருவாகின்றன. இந்த பைட்டோ கெமிக்கல்கள் இன்று பயன்படுத்தப்படும் Nrf2-செயல்படுத்தும் மருந்து தயாரிப்புகளைப் போலவே குறைவான பக்க விளைவுகளுடன் சமமாக சக்தி வாய்ந்ததாகத் தெரிகிறது.
நியூக்ளியர் காரணி எரித்ராய்டு 2-தொடர்புடைய காரணி, பொதுவாக Nrf2 என அழைக்கப்படுகிறது, இது மரபணுக்கள், நொதிகள் மற்றும் ஆக்ஸிஜனேற்ற பதில்களை ஒழுங்குபடுத்துவதன் மூலம் செல்லைப் பாதுகாக்கும் ஒரு டிரான்ஸ்கிரிப்ஷன் காரணியாகும். டிரான்ஸ்கிரிப்ஷன் காரணிகள் என்பது ஒரு வகை புரதமாகும், இது குளுதாதயோன் எஸ்-டிரான்ஸ்ஃபெரேஸ்கள் அல்லது ஜிஎஸ்டிகள் உட்பட குறிப்பிட்ட பொருட்கள் மற்றும் இரசாயனங்களை உருவாக்குவதை ஊக்குவிக்க டிஎன்ஏவுடன் இணைக்கப்பட்டுள்ளது. Nrf2 செயல்படுத்தல் செயலில் உள்ள புரதங்களின் உற்பத்தியைத் தூண்டுகிறது, இது ஆக்ஸிஜனேற்ற அழுத்தத்தைக் குறைக்க உதவும் சக்திவாய்ந்த ஆக்ஸிஜனேற்ற திறனை வெளிப்படுத்துகிறது.
டாக்டர். அலெக்ஸ் ஜிமினெஸ் DC, CCST இன்சைட்
Nrf2 செயல்படுத்தலுக்குப் பின்னால் உள்ள அறிவியல்
2 இல் ஆரம்ப Nrf2004-செயல்படுத்தும் உணவு நிரப்பி உருவாக்கப்பட்டவுடன், Nrf2 பாதையின் செயல்பாடு குறித்து குறைந்தபட்ச தகவல் அறியப்பட்டது. Nrf200 பற்றிய இலக்கியத்தில் சுமார் 2 செய்தித்தாள்கள், அணுக்கரு காரணி போன்ற 2 அல்லது NFE2L2 என்றும் அழைக்கப்படுகின்றன, மேலும் ஆராய்ச்சியாளர்கள் பாலூட்டிகளில் Nrf2 இன் ஆக்ஸிஜனேற்ற பதிலைக் கண்டறியத் தொடங்கினர். இருப்பினும், 2017 ஆம் ஆண்டு வரை, இந்த "மாஸ்டர் ரெகுலேட்டர்" குறித்த 9,300 க்கும் மேற்பட்ட கல்வி ஆராய்ச்சி ஆய்வுகள் அச்சிடப்பட்டுள்ளன.
உண்மையில், Nrf2 மரபணுக்களுடன் தொடர்புபடுத்தாத பல ஆக்ஸிஜனேற்ற நொதிகளை ஒழுங்குபடுத்துகிறது, மாறாக, ஆரோக்கியமான மற்றும் நோயியல் நிலைமைகளின் கீழ் செல்கள், உறுப்புகள் மற்றும் இறுதியில் உயிரினங்கள் எதிர்கொள்ளும் மன அழுத்தம் தொடர்பான பல்வேறு சூழ்நிலைகளுக்கு எதிராக அவை பாதுகாப்பை வழங்குகின்றன. வெளியிடப்பட்ட கல்வி ஆராய்ச்சி ஆய்வுகளின் இந்த புதிய அளவிலான தகவல்களின் அடிப்படையில், ஆராய்ச்சியாளர்கள் இப்போது சிறப்பாக உருவாக்க முடியும் Nrf2 உணவு சப்ளிமெண்ட்ஸ்.
2007 ஆம் ஆண்டு வரை, ஆராய்ச்சி ஆய்வுகள் Nrf2 பாதையின் சிக்கலான செயல்பாட்டை நிரூபித்துள்ளன. Nrf2 ஆக்டிவேட்டர்கள் மனித உடலில் உள்ள பல்வேறு கட்டமைப்புகளின் காரணிகளைப் பிரதிபலிக்கின்றன. இந்த பாதைகள் மூலம், Nrf2 ஆக்டிவேட்டர்கள், சமநிலையை வைத்து, மரபணுக்களின் தேவைகளுக்குப் பதிலளிக்கும் வகையில், செல் முழுவதும் மாறிவரும் நிலைமைகளை உணரும் வகையில் பொருத்தப்பட்டுள்ளன.
ஏன் Nrf2-செயல்படுத்தும் சப்ளிமெண்ட்ஸ் பயன்படுத்த வேண்டும்?
Nrf2-செயல்படுத்தும் திறன்கள் உயிரினங்களில் வயதுக்கு ஏற்ப குறைவதால், மாற்றங்கள் ஏற்பட ஆரம்பிக்கலாம். உயிரணுக்களில் Nrf2 இன் கவனம் வயதுக்கு ஏற்ப குறைகிறது, ஆக்ஸிஜனேற்ற அழுத்தத்தின் அதிகரித்த குறிப்பான்களைக் காட்டுகிறது என்பதை ஆராய்ச்சி ஆய்வுகள் நிரூபித்துள்ளன. பெருந்தமனி தடிப்பு மற்றும் இருதய நோய், மூட்டுவலி, புற்றுநோய், உடல் பருமன், வகை 2 நீரிழிவு, உயர் இரத்த அழுத்தம், கண்புரை மற்றும் அல்சைமர் நோய் மற்றும் பார்கின்சன் நோய் போன்ற பல்வேறு வயது தொடர்பான நோய்கள் இந்த மாற்றங்களால் உருவாகலாம். இந்த உடல்நலப் பிரச்சினைகளுடன் ஆக்ஸிஜனேற்ற அழுத்தம் கண்டறியப்பட்டுள்ளது.
Nrf2 ஆக்டிவேட்டர்களின் உற்பத்தியை அதிகரிக்க செல்லின் திறனை தூண்டுவதன் மூலம், Nrf2 உணவு சப்ளிமெண்ட்ஸ் ஆக்ஸிஜனேற்ற அழுத்தத்தின் விளைவுகளை எதிர்க்கும் மனித உடலின் சொந்த திறனை புதுப்பிக்க உதவும். பாலிஅன்சாச்சுரேட்டட் கொழுப்பு அமிலங்கள், அல்லது PUFAகள், மிகவும் எளிதில் ஆக்ஸிஜனேற்றப்பட்ட மூலக்கூறுகளில் ஒன்றாகும், மேலும் அவை குறிப்பாக ஃப்ரீ ரேடிக்கல்களால் பாதிக்கப்படக்கூடியவை. தியோபார்பிட்யூரிக் அமிலம், அல்லது TBARS, உற்பத்தியானது வயதுக்கு ஏற்ப அதிகரிக்கலாம், இது Nrf2-ஒழுங்குபடுத்தப்பட்ட பாதைகளில் வீழ்ச்சியுடன் கூடிய ஆக்ஸிஜனேற்ற அழுத்தத்தைக் குறிக்கிறது.
உயிரியல் ரீதியாக, மரபணு தூண்டல் என்பது மிகவும் மெதுவான பொறிமுறையாகும், பொதுவாக ஒரு பாதை வழியாக மாற்றுவதற்கு மணிநேரம் தேவைப்படுகிறது. இதன் விளைவாக, பல நொதிகள் அவற்றின் சொந்த ஆன்/ஆஃப் சுவிட்சுகளைக் கொண்டுள்ளன, அவை வெவ்வேறு ஒழுங்குமுறை நொதிகளால் நிமிடங்களில் தூண்டப்படலாம். ஆராய்ச்சியாளர்கள் Nrf2 ஆக்டிவேட்டர்களின் தனியுரிம கலவைகளை உருவாக்கியுள்ளனர், இது இந்த அறிவுத் தளத்தை செயல்படுத்துகிறது. Nrf2 செயல்படுத்தல் ஆனது Nrf2 டிரான்ஸ்கிரிப்ஷன் காரணி அதன் தடுப்பானிலிருந்து வெளியேற்றப்பட்டு செல் அணுக்கருவிற்கு இடம்பெயர்வதோடு மட்டுமல்லாமல், சைட்டோபுரோடெக்டிவ் மரபணு வெளிப்பாட்டைத் தூண்டுவதற்கு குறிப்பிட்ட DNA வரிசைகளுடன் பிணைக்கப்பட்டு, Nrf2 கருவில் இருந்து எடுக்கப்படும் வேகத்தை ஒழுங்குபடுத்துகிறது.
நீக்குதல் செயல்முறை மற்றும் மனித உடலில் Nrf2 செயல்படுத்துதல் ஆகியவற்றைப் புரிந்துகொள்வது, மரபணுக்களின் பிரதிபலிப்பை அதன் பண்பேற்றம் மூலம் நிறைவேற்ற பல்வேறு Nrf2 ஆக்டிவேட்டர்களின் சேர்க்கைகளை உருவாக்க ஆராய்ச்சியாளர்களை அனுமதித்துள்ளது. அறிவுத் தளத்தின் கலவையானது, பல்வேறு வகையான பிற ஆராய்ச்சி ஆய்வுகளுடன் சேர்ந்து, உணவுப் பொருட்களாகப் பயன்படுத்த Nrf2 ஆக்டிவேட்டர்களை உருவாக்க உதவியது. எங்கள் தகவலின் நோக்கம் உடலியக்க மற்றும் முதுகெலும்பு சுகாதார பிரச்சினைகள் மட்டுமே. விஷயத்தைப் பற்றி விவாதிக்க, தயவு செய்து டாக்டர் ஜிமினெஸைக் கேட்கவும் அல்லது எங்களை இங்கே தொடர்பு கொள்ளவும்915-850-0900.
டாக்டர் அலெக்ஸ் ஜிமெனெஸ் தொகுத்தார்
கூடுதல் தலைப்பு விவாதம்: அறுவை சிகிச்சை இல்லாமல் முழங்கால் வலியை நீக்குதல்
முழங்கால் வலி என்பது நன்கு அறியப்பட்ட அறிகுறியாகும், இது பல்வேறு முழங்கால் காயங்கள் மற்றும்/அல்லது நிலைமைகள் காரணமாக ஏற்படலாம்.விளையாட்டு காயங்கள். முழங்கால் மனித உடலில் மிகவும் சிக்கலான மூட்டுகளில் ஒன்றாகும், ஏனெனில் இது நான்கு எலும்புகள், நான்கு தசைநார்கள், பல்வேறு தசைநாண்கள், இரண்டு மெனிசிஸ் மற்றும் குருத்தெலும்பு ஆகியவற்றின் குறுக்குவெட்டுகளால் ஆனது. அமெரிக்கன் அகாடமி ஆஃப் ஃபேமிலி பிசிஷியன்களின் கூற்றுப்படி, முழங்கால் வலிக்கான பொதுவான காரணங்களில் பட்டேலர் சப்லக்சேஷன், பட்டேலர் டெண்டினிடிஸ் அல்லது ஜம்பர்ஸ் முழங்கால் மற்றும் ஓஸ்குட்-ஸ்க்லாட்டர் நோய் ஆகியவை அடங்கும். முழங்கால் வலி 60 வயதுக்கு மேற்பட்டவர்களுக்கு ஏற்பட வாய்ப்புள்ளது என்றாலும், குழந்தைகள் மற்றும் இளம்பருவத்தினருக்கும் முழங்கால் வலி ஏற்படலாம். முழங்கால் வலியை RICE முறைகளைப் பின்பற்றி வீட்டிலேயே சிகிச்சை செய்யலாம், இருப்பினும், கடுமையான முழங்கால் காயங்களுக்கு உடலியக்க சிகிச்சை உட்பட உடனடி மருத்துவ கவனிப்பு தேவைப்படலாம்.
ஆன்டிஆக்ஸிடன்ட்கள் மனித உடலில் ஆக்சிஜனேற்ற செயல்முறையை கட்டுப்படுத்தும் சேர்மங்கள் என்று அறிவியல் பூர்வமாக குறிப்பிடப்படுகின்றன, அவை சரிபார்க்கப்படாமல் விட்டால், செல்லுலார் சேதத்தை ஏற்படுத்தக்கூடிய பல சங்கிலி எதிர்வினைகளை உருவாக்கக்கூடிய ஃப்ரீ ரேடிக்கல்களை உருவாக்கலாம். அதிர்ஷ்டவசமாக, மனித உடலால் அத்தகைய உள்ளமைக்கப்பட்ட நோயெதிர்ப்பு வழிமுறைகளை உருவாக்க முடியும், இருப்பினும், வினைத்திறன் ஆக்ஸிஜன் இனங்கள் அல்லது ROS ஐ நடுநிலையாக்க முடியாமல் போகும் போது, ஆக்ஸிஜனை உட்செலுத்தும்போது கட்டுப்பாட்டை மீறும் ஒரு சிறிய சுடரை கற்பனை செய்து, தீங்கு நேரிடும். .
சுடரின் உருவகத்தை விரிவுபடுத்துவதைத் தொடர, ROS அல்லது எதிர்வினை ஆக்ஸிஜன் இனங்களின் தாக்கத்தை நடுநிலையாக்கும் திறன் இல்லாததன் இறுதி விளைவு சேதம் மற்றும் வீக்கம் ஆகும், வேறுவிதமாகக் கூறினால், மனித உடல் உண்மையில் தீயில் எரிகிறது. அற்புதமான விஷயம் என்னவென்றால், இந்த உடல்நலப் பிரச்சினையை எதிர்த்துப் போராட உதவும் ஆன்டிஆக்ஸிடன்ட்கள் உள்ளன, மேலும் இந்த ஆன்டிஆக்ஸிடன்ட் குளுதாதயோன் ஆகும். 1889 இல் கண்டுபிடிக்கப்பட்டாலும், குளுதாதயோனின் ஆக்ஸிஜனேற்ற விளைவு நவீன ஆராய்ச்சி ஆய்வுகளில் மிகவும் சுவாரஸ்யமான தலைப்புகளில் ஒன்றாக மாறியுள்ளது.
ஆன்டிஆக்ஸிடன்ட்களின் மாஸ்டர்: குளுதாதயோன்
சக்தி வாய்ந்த பொருள் சிஸ்டைன், குளுடாமிக் அமிலம் மற்றும் கிளைசின் ஆகியவற்றிலிருந்து உருவாகும் டிரிபெப்டைட் ஆகும். ஃப்ரீ ரேடிக்கல்களின் உருவாக்கத்திற்கு எதிராக மனித உடலைப் பாதுகாக்கும் அதன் திறன் காரணமாக, குளுதாதயோன் இறுதியில் ஆரோக்கியமான நோயெதிர்ப்பு மண்டலத்தை மேம்படுத்த உதவுகிறது. அடிப்படையில் 2015 இல் அறிவியல் அறிக்கைகள், பெராக்ஸிரெடின் மற்றும் கேடலேஸுடன் இணைந்து செயல்படும் குளுதாதயோனின் திறன் ஹைட்ரஜன் பெராக்சைடுக்கு எதிராக செல்களைப் பாதுகாக்க உதவுகிறது என்று தீர்மானிக்கப்பட்டது. இந்த சினெர்ஜிஸ்டிக் ஃபார்முலா எதிர்வினை ஆக்ஸிஜன் இனங்கள் அல்லது ROS க்கு எதிராக செயல்படுகிறது. குளுதாதயோன், பெராக்சிட்ரெடின் மற்றும் கேடலேஸ் ஆகியவை செல்லுலார் ஹோமியோஸ்டாசிஸின் அதிகரிப்புக்கு இன்றியமையாத கூறுகள் ஆகும், இது ஆரோக்கியமான செல்கள், திசுக்கள் மற்றும் உறுப்புகளின் இன்றியமையாத செயல்முறையாகும்.
கூடுதலாக, குளுதாதயோன் லிம்போசைட் செயல்பாடுகளில் அதன் முக்கிய விளைவைப் பயன்படுத்தி ஒட்டுமொத்த நோயெதிர்ப்பு அமைப்பு மற்றும் செயல்பாட்டை அதிகரிக்கிறது. அதில் கூறியபடி இம்யூனோ கெமிஸ்ட்ரி துறை, மனித உடலில் குளுதாதயோனின் அளவை சரியாக நிரப்புவது நோயெதிர்ப்பு எதிர்வினைகளை பெரிதும் மேம்படுத்தும். உதாரணமாக, இரண்டு சீரற்ற மருந்துப்போலி கட்டுப்படுத்தப்பட்ட சோதனைகள், N-acetyl-cysteine அல்லது NAC உடன் நோயெதிர்ப்பு-சமரசம் செய்யப்பட்ட நோயாளிகளின் சிகிச்சை சிகிச்சையின் விளைவாக, இரண்டு நிகழ்வுகளிலும், முழு புத்துணர்ச்சியையும் உள்ளடக்கிய பெரும்பாலான நோயெதிர்ப்பு செயல்முறைகளில் கணிசமான வளர்ச்சி ஏற்பட்டது. இயற்கை கொலையாளி செல் செயல்பாடு. N-acetyl-cysteine, அல்லது NAC, குளுதாதயோனில் இருந்து கந்தகத்தைப் பயன்படுத்துகிறது மற்றும் அதை விஷ மூலக்கூறுகளுடன் இணைக்கிறது, பின்னர் அவை நீரில் கரையக்கூடியதாக மாறி மனித உடலில் வெளியேற்றப்படுகின்றன.
க்ளூதாதயோனுக்கு லிபோயிக் அமிலத்தை புத்துயிர் அளிப்பதற்கும், வைட்டமின் சி மற்றும் ஈ ஆகியவற்றை மறுசுழற்சி செய்யும் திறனும் உள்ளது, இது ஃப்ரீ ரேடிக்கல்களை நடுநிலையாக்க எலக்ட்ரான்களை அனுப்புவதன் மூலம் சில அமைப்பு செயல்முறைகளைத் தொடங்குவதற்குத் தேவையானது. ஒரு ஆராய்ச்சி ஆய்வின் அடிப்படையில் PLOS ONE, குளுதாதயோன் நீரிழிவு மெட்டிலஸ், அல்லது T2DM மற்றும் மைக்கோபாக்டீரியம் காசநோயால் பாதிக்கப்பட்ட நோயாளிகள். பொதுவாக, பலவீனமான நோயெதிர்ப்பு அமைப்பு கொண்ட நபர்கள் M. tb, அல்லது மைக்கோபாக்டீரியம் காசநோய், நோய் அல்லது தொற்றுக்கு அதிக வெளிப்பாட்டைக் காண்பிக்கும் போக்கைக் கொண்டுள்ளனர். மேலும், T2DM இல்லாதவர்களைக் காட்டிலும் T2DM அல்லது T2DM உள்ளவர்கள் TB நோயால் பாதிக்கப்படுவதற்கான வாய்ப்புகள் இரண்டு முதல் மூன்று மடங்கு அதிகம். T2DM உள்ள நோயாளிகளிடமிருந்து தனிமைப்படுத்தப்பட்ட மேக்ரோபேஜ்களில் குளுதாதயோனின் அளவை அதிகரிப்பது M.Tb நோய் அல்லது நோய்த்தொற்றின் மேம்பட்ட கட்டுப்பாட்டிற்கு வழிவகுத்தது என்றும் ஆராய்ச்சி ஆய்வு பரிந்துரைத்தது. T2DM உள்ள நோயாளிகளில் குளுதாதயோனின் குறைந்த அளவு M. tb நோய் அல்லது நோய்த்தொற்றின் அதிக வாய்ப்புக்கு பங்களிக்கிறது என்பதை இந்த முடிவுகள் நிரூபிக்கின்றன. மேலும், சார்ந்தது பிரைட்டன் மற்றும் சசெக்ஸ் மருத்துவப் பள்ளியில் டீட்ரோ கெஸ்ஸி, ஆக்ஸிஜனேற்ற அழுத்தம் இறுதியில் மோசமான நோயெதிர்ப்பு அமைப்பு அமைப்பு மற்றும் செயல்பாட்டை ஏற்படுத்தும்.
அதிர்ஷ்டவசமாக, குளுதாதயோன் நோய் எதிர்ப்பு சக்தியை வலுப்படுத்துவதிலும் கட்டுப்படுத்துவதிலும் முக்கிய பங்கு வகிக்கிறது. உதாரணமாக, டி-லிம்போசைட் பெருக்கம், பாலிமார்போநியூக்ளியர் நியூட்ரோபில்களின் பாகோசைடிக் செயல்பாடு மற்றும் டென்ட்ரிடிக் செல் செயல்பாடுகள் உள்ளிட்ட நோயெதிர்ப்பு மண்டலத்தில் உள்ள உள்ளார்ந்த மற்றும் தழுவல் செயல்முறைகளுக்கு குளுதாதயோன் இன்றியமையாதது. . செல்-தியானம் செய்யப்பட்ட நோய் எதிர்ப்பு சக்தியில் புரத ஆன்டிஜென்கள் அடங்கும், அவை ஆரம்பத்தில் மேக்ரோபேஜ்கள் மற்றும் டென்ட்ரிடிக் செல்களின் எண்டோசைடிக் வெசிகிள்களில் சிதைவடையத் தொடங்குகின்றன, எனவே, ஆன்டிஜென்-குறிப்பிட்ட T செல்களின் பெருக்கத்தை செயல்படுத்த சிறிய பெப்டைடுகள் மேற்பரப்பில் நிரூபிக்கப்படுகின்றன. கூடுதலாக, குளுதாதயோன் சைட்டோகைன்களை உருவாக்க உதவுகிறது, மேலும் டென்ட்ரிடிக் செல்கள் மூலம் இன்டர்ஃபெரான்-காமா உற்பத்தியை பராமரிப்பது அவசியம், இது மைக்கோபாக்டீரியா உள்ளிட்ட உள்செல்லுலார் நோய்க்கிருமிகளுக்கு எதிராக பாதுகாப்பதில் முக்கியமானது.
N-acetyl-cysteine, அல்லது NAC, அறிவியல் ரீதியாக குளுதாதயோனின் முன்னோடி என குறிப்பிடப்படுகிறது, மேலும் இது மிகவும் சக்திவாய்ந்த செல்லுலார் ஆக்ஸிஜனேற்றம் ஆகும், இது ஒரு ஃப்ரீ ரேடிக்கல் ஸ்கேவெஞ்சர் ஆக்ஸிஜனேற்றியாகப் பயன்படுத்தப்படுகிறது. அதன் பங்கிற்காக பொதுவாக அங்கீகரிக்கப்பட்டது அசிடமினோஃபென் நச்சுத்தன்மையைத் தடுக்கிறது, NAC, or'N-acetyl-cysteine, பல உடல்நலம் மற்றும் ஆரோக்கிய நன்மைகளைக் கொண்டிருப்பதாக நிரூபிக்கப்பட்டுள்ளது. படி செல் ஜர்னல், NAC ஒரு ஆரோக்கியமான அழற்சி பதிலை ஆதரிக்க உதவுகிறது மற்றும் மனித கால மற்றும் முன்கூட்டிய பிரசவத்தை சாதகமாக பாதிக்கலாம். முந்தைய குறைப்பிரசவம் மற்றும் பாக்டீரியா வஜினோசிஸ் உள்ள பெண்களில், கர்ப்பத்தின் 0.6 வது வாரத்திற்குப் பிறகு புரோஜெஸ்ட்டிரோனுடன் வாய்வழியாக எடுத்துக் கொள்ளப்பட்ட ஒரு நாளைக்கு 16 கிராம் என்ஏசி, குறைப்பிரசவம் மற்றும் மேம்படுத்தப்பட்ட குழந்தை பிறப்பிலிருந்து பாதுகாக்கிறது என்று ஆராய்ச்சி ஆய்வு முடிவு செய்தது. முடிவில், தசைக் கட்டமைப்பில் NAC இன் நேர்மறையான விளைவுகளும் கண்டறியப்பட்டன. மூன்று நிமிட தொடர்ச்சியான சுருக்கங்களுக்குப் பிறகு, 15 சதவிகிதம் மேம்பட்ட வெளியீடு இருந்தது, தசைக் கட்டமைப்பை மேம்படுத்துவதிலும், பிரசவத்தின் போது ஒட்டுமொத்த சோர்வைக் குறைப்பதிலும் NAC எவ்வாறு அடிப்படைப் பங்கு வகிக்கிறது என்பதை நிரூபிக்கிறது.
NAC, or'N-acetyl-cysteine, பாலிசிஸ்டிக் ஓவரியன் சிண்ட்ரோம் அல்லது பிசிஓஎஸ் உள்ளவர்களுக்கு பயனளிக்கும் என்பதையும் ஆராய்ச்சியாளர்கள் கண்டுபிடித்துள்ளனர். பிசிஓஎஸ், அல்லது பாலிசிஸ்டிக் ஓவேரியன் சிண்ட்ரோம், இது ஒரு பொதுவான நாளமில்லா சுரப்பிகள் தொடர்பான நோயாகும், இது இனப்பெருக்க வயதுடைய பெண்களில் சுமார் 5 முதல் 10 சதவீதத்தை பாதிக்கிறது. அத்தகைய நோயாளிகளில், மெட்டபாலிக் சிண்ட்ரோம் ஏற்படுவதற்கான அதிக ஆபத்து உள்ளது, அங்கு NAC இன் பயன்பாடு ஆரோக்கியமான இன்சுலின் அளவையும் உணர்திறனையும் மீட்டெடுக்க உதவியது.
டாக்டர் அலெக்ஸ் ஜிமெனெஸின் நுண்ணறிவு
ஒட்டுமொத்த ஆரோக்கியத்தையும் ஆரோக்கியத்தையும் அடைவதிலும் பராமரிப்பதிலும் அதன் அடிப்படைப் பங்கு காரணமாக குளுதாதயோன் "ஆன்டிஆக்ஸிடன்ட்களின் மாஸ்டர்" என்று குறிப்பிடப்படுகிறது. மனித உடல் அதன் சொந்த குளுதாதயோனை உற்பத்தி செய்யும் திறன் கொண்டது, மோசமான ஊட்டச்சத்து, மாசுபாடு, நச்சுகள், மருந்துகள் மற்றும்/அல்லது மருந்துகளின் அதிகப்படியான பயன்பாடு, மன அழுத்தம், அதிர்ச்சி, முதுமை, நோய் மற்றும் கதிர்வீச்சு ஆகியவை குளுதாதயோனின் இயற்கையான அளவைக் குறைக்கலாம். இது ஆக்ஸிஜனேற்ற அழுத்தம், ஃப்ரீ ரேடிக்கல்கள், நோய்த்தொற்றுகள் மற்றும் புற்றுநோய் ஆகியவற்றிலிருந்து செல் சேதத்திற்கு தனிநபர்களை மிகவும் எளிதில் பாதிக்கலாம். எனவே குளுதாதயோன் சப்ளிமெண்ட்ஸ் மனித உடலில் மிகப்பெரிய நன்மைகளை ஏற்படுத்தும். உடலியக்க சிகிச்சை போன்ற மாற்று சிகிச்சை விருப்பங்களுடன் சேர்ந்து, குளுதாதயோன் அளவை மீண்டும் ஒருமுறை நல்வாழ்வை மேம்படுத்த ஒழுங்குபடுத்தலாம்.
கூடுதலாக, சுகாதார வல்லுநர்கள் மற்ற மாற்று சிகிச்சை விருப்பங்களுடன் குளுதாதயோன் கூடுதல் பயன்பாட்டை செயல்படுத்த பரிந்துரைத்துள்ளனர். உடலியக்க சிகிச்சை, ஒட்டுமொத்த ஆரோக்கியத்தையும் ஆரோக்கியத்தையும் மேலும் மேம்படுத்த. ஆக்ஸிஜனேற்றிகள் அதிகபட்ச நல்வாழ்வை பராமரிப்பதற்கும், செல் தீங்கு அல்லது சேதத்தை ஏற்படுத்தும் ஃப்ரீ ரேடிக்கல்களின் சங்கிலி எதிர்வினையைத் தடுப்பதற்கும் முக்கியம். மேலே குறிப்பிட்டுள்ளபடி, குளுதாதயோன் போன்ற சக்திவாய்ந்த ஆக்ஸிஜனேற்றிகள் இறுதியில் இந்த ஃப்ரீ ரேடிக்கல்களின் வளர்ச்சியைக் கட்டுப்படுத்த உதவுகின்றன மற்றும் ஆரோக்கியமான நோயெதிர்ப்பு மண்டலத்தை வழங்குகின்றன. என்று ஆய்வுகள் கண்டறிந்துள்ளன உடலியக்க சிகிச்சை இந்த செயல்பாட்டில் இன்றியமையாத பங்கு வகிக்கலாம், இயற்கையாகவே மனித உடலில் ஆக்ஸிஜனேற்றத்தின் செயல்பாட்டை அதிகரிக்கும். சிரோபிராக்டிக் கவனிப்பு என்பது பாதுகாப்பான மற்றும் பயனுள்ள சிகிச்சை அணுகுமுறையாகும், இது மருந்துகள்/மருந்துகள் மற்றும்/அல்லது அறுவை சிகிச்சை தலையீடுகள் இல்லாமல் மனித உடலை இயற்கையாகவே குணப்படுத்த அனுமதிக்கும் வகையில், முதுகெலும்பு சீரமைப்புகள் மற்றும் கையேடு கையாளுதல்களை முதுகுத்தண்டின் தவறான சீரமைப்புகள் அல்லது சப்லக்சேஷன்களை சரி செய்ய பயன்படுத்துகிறது.
இறுதியாக, ஆன்டிஆக்ஸிடன்ட்கள் தங்கள் உயிரியல் பண்புகளை ஏராளமான ஆரோக்கிய நன்மைகள் மூலம் வெளிப்படுத்துகின்றன, இது நமது நவீன உலகில் அதிகரித்து வரும் மன அழுத்தம், நோய் மற்றும் மாசுபாட்டின் ஒவ்வொரு தாக்குதலுக்கும் அவசியமாக இருக்கலாம், இவை அனைத்தும் செல் தீங்கு மற்றும்/அல்லது சேதத்திற்கு பங்களிக்கின்றன. . குளுதாதயோன் மற்றும் அதன் முன்னோடி, NAC, or'N-acetyl-cysteine, ஆன்ட்டி ஆக்சிடென்ட்களின் மண்டலத்தில் அவற்றின் சக்திவாய்ந்த நிலையைத் தொடர்ந்து காட்டுகின்றன. உடலியக்க சிகிச்சை போன்ற மாற்று சிகிச்சை விருப்பங்களுடன் சேர்ந்து, இந்த சக்திவாய்ந்த ஆக்ஸிஜனேற்றம் வழங்கும் அனைத்து நன்மைகளையும் மக்கள் பயன்படுத்திக் கொள்ளலாம். எங்கள் தகவலின் நோக்கம் உடலியக்க சிகிச்சை மற்றும் முதுகெலும்பு காயங்கள் மற்றும் நிலைமைகளுக்கு மட்டுமே. விஷயத்தைப் பற்றி விவாதிக்க, தயவு செய்து டாக்டர் ஜிமினெஸைக் கேட்கவும் அல்லது எங்களை இங்கே தொடர்பு கொள்ளவும்915-850-0900.
டாக்டர் அலெக்ஸ் ஜிமெனெஸ் தொகுத்தார்
கூடுதல் தலைப்புகள்: முதுகுவலி
முதுகு வலி இயலாமைக்கான மிகவும் பிரபலமான காரணங்களில் ஒன்றாகும் மற்றும் உலகளவில் வேலையில் தவறவிட்ட நாட்கள். உண்மையில், முதுகுவலி என்பது மருத்துவர் அலுவலக வருகைகளுக்கான இரண்டாவது பொதுவான காரணமாகக் கூறப்படுகிறது, இது மேல் சுவாச நோய்த்தொற்றுகளால் மட்டுமே. ஏறக்குறைய 80 சதவிகித மக்கள் தங்கள் வாழ்நாள் முழுவதும் ஒரு முறையாவது சில வகையான முதுகுவலியை அனுபவிப்பார்கள். முதுகெலும்பு என்பது எலும்புகள், மூட்டுகள், தசைநார்கள் மற்றும் தசைகள் ஆகியவற்றால் ஆன ஒரு சிக்கலான கட்டமைப்பாகும். இதன் காரணமாக, காயங்கள் மற்றும் / அல்லது மோசமான நிலைமைகள் போன்றவை ஹெர்னியேட்டட் டிஸ்க்குகள், இறுதியில் முதுகுவலியின் அறிகுறிகளுக்கு வழிவகுக்கும். விளையாட்டு காயங்கள் அல்லது ஆட்டோமொபைல் விபத்து காயங்கள் பெரும்பாலும் முதுகுவலிக்கு அடிக்கடி காரணமாகின்றன, இருப்பினும், சில நேரங்களில் எளிமையான இயக்கங்கள் வலிமிகுந்த முடிவுகளை ஏற்படுத்தும். அதிர்ஷ்டவசமாக, சிரோபிராக்டிக் பராமரிப்பு போன்ற மாற்று சிகிச்சை விருப்பங்கள், முதுகெலும்பு சரிசெய்தல் மற்றும் கையேடு கையாளுதல்களைப் பயன்படுத்துவதன் மூலம் முதுகுவலியைக் குறைக்க உதவும், இறுதியில் வலி நிவாரணத்தை மேம்படுத்துகின்றன.
அறிவியல் அடிப்படையிலான சிரோபிராக்டர் டாக்டர். அலெக்சாண்டர் ஜிமினெஸ் பாருங்கள் விஷத்தன்மை அழுத்தம், அது என்ன, அது உடலை எவ்வாறு பாதிக்கிறது மற்றும் நிலைமையை சரிசெய்ய ஆக்ஸிஜனேற்ற பாதுகாப்பு.
எஸ்ரா பிர்பென் PhD, 1 Umit Murat Sahiner MD, 1 Cansin Sackesen MD, 1 Serpil Erzurum MD,2 மற்றும் Omer Kalayci, MD1
சுருக்கம்: எதிர்வினை ஆக்ஸிஜன் இனங்கள் (ROS) சாதாரண செல்லுலார் வளர்சிதை மாற்றம் மற்றும் சுற்றுச்சூழல் காரணிகளான காற்று மாசுபாடுகள் அல்லது சிகரெட் புகை போன்றவற்றின் விளைவாக உயிரினங்களால் உற்பத்தி செய்யப்படுகின்றன. ROS மிகவும் எதிர்வினை மூலக்கூறுகள் மற்றும் கார்போஹைட்ரேட்டுகள், நியூக்ளிக் அமிலங்கள், லிப்பிடுகள் மற்றும் புரதங்கள் போன்ற செல் கட்டமைப்புகளை சேதப்படுத்தும் மற்றும் அவற்றின் செயல்பாடுகளை மாற்றும். ஆக்சிடென்ட்களுக்கு ஆதரவாக ஆக்சிடென்ட்கள் மற்றும் ஆன்டிஆக்ஸிடன்ட்டுகளுக்கு இடையே உள்ள சமநிலையில் ஏற்படும் மாற்றத்தை ஆக்ஸிஜனேற்ற அழுத்தம் என்று அழைக்கப்படுகிறது. ஏரோபிக் உயிரினங்கள் ஒருங்கிணைந்த ஆக்ஸிஜனேற்ற அமைப்புகளைக் கொண்டுள்ளன, இதில் நொதி மற்றும் நொதி அல்லாத ஆக்ஸிஜனேற்றங்கள் பொதுவாக ROS இன் தீங்கு விளைவிக்கும் விளைவுகளைத் தடுப்பதில் பயனுள்ளதாக இருக்கும். இருப்பினும், நோயியல் நிலைகளில், ஆக்ஸிஜனேற்ற அமைப்புகள் அதிகமாக இருக்கலாம். ஆக்ஸிஜனேற்ற அழுத்தமானது புற்றுநோய், நரம்பியல் கோளாறுகள், பெருந்தமனி தடிப்பு, உயர் இரத்த அழுத்தம், இஸ்கிமியா/பெர்ஃப்யூஷன், நீரிழிவு, கடுமையான சுவாசக் கோளாறு நோய்க்குறி, இடியோபாடிக் நுரையீரல் ஃபைப்ரோஸிஸ், நாள்பட்ட தடுப்பு நுரையீரல் நோய் மற்றும் ஆஸ்துமா உள்ளிட்ட பல நோயியல் நிலைமைகள் மற்றும் நோய்களுக்கு பங்களிக்கிறது. இந்த மதிப்பாய்வில், செல்லுலார் ஆக்ஸிஜனேற்ற மற்றும் ஆக்ஸிஜனேற்ற அமைப்புகளை நாங்கள் சுருக்கமாகக் கூறுகிறோம் மற்றும் ஆக்ஸிஜனேற்ற அழுத்தத்தின் செல்லுலார் விளைவுகள் மற்றும் வழிமுறைகளைப் பற்றி விவாதிக்கிறோம்.
முக்கிய வார்த்தைகள்: ஆக்ஸிஜனேற்ற, ஆக்ஸிஜனேற்ற, ஆக்ஸிஜனேற்ற அழுத்தம், எதிர்வினை ஆக்ஸிஜன் இனங்கள், ரெடாக்ஸ்
(WAO ஜர்னல் 2012; 5:9-19)
எதிர்வினை ஆக்ஸிஜன் இனங்கள் (ROS) சாதாரண செல்லுலார் வளர்சிதை மாற்றத்தின் விளைவாக உயிரினங்களால் உற்பத்தி செய்யப்படுகின்றன. குறைந்த முதல் மிதமான செறிவுகளில், அவை உடலியல் உயிரணு செயல்முறைகளில் செயல்படுகின்றன, ஆனால் அதிக செறிவுகளில், அவை லிப்பிடுகள், புரதங்கள் மற்றும் டிஎன்ஏ போன்ற செல் கூறுகளுக்கு பாதகமான மாற்றங்களை உருவாக்குகின்றன. ஆக்ஸிஜனேற்ற அழுத்தம் என்று அழைக்கப்படுகிறது ,1 மற்றும் ஆஸ்துமா.6–7 ஏரோபிக் உயிரினங்கள் ஒருங்கிணைந்த ஆக்ஸிஜனேற்ற அமைப்புகளைக் கொண்டுள்ளன, இதில் நொதி மற்றும் நொதி அல்லாத ஆக்ஸிஜனேற்றங்கள் உள்ளன, அவை பொதுவாக ROS இன் தீங்கு விளைவிக்கும் விளைவுகளைத் தடுப்பதில் பயனுள்ளதாக இருக்கும். இருப்பினும், நோயியல் நிலைகளில், ஆக்ஸிஜனேற்ற அமைப்புகள் அதிகமாக இருக்கலாம். இந்த மதிப்பாய்வில், செல்லுலார் ஆக்சிடென்ட் மற்றும் ஆக்ஸிஜனேற்ற அமைப்புகள் மற்றும் ஆரோக்கியம் மற்றும் நோய் நிலைகளில் குறைக்கும் மற்றும் ஆக்சிஜனேற்றம் (ரெடாக்ஸ்) நிலையை ஒழுங்குபடுத்துதல் ஆகியவற்றை சுருக்கமாகக் கூறுகிறோம்.
ஆக்ஸிஜனேற்றங்கள்
ROS இன் எண்டோஜெனஸ் ஆதாரங்கள்
ROS சாதாரண செல்லுலார் வளர்சிதை மாற்றத்தின் விளைவாக மூலக்கூறு ஆக்ஸிஜனில் இருந்து தயாரிக்கப்படுகிறது. ROS ஐ 2 குழுக்களாகப் பிரிக்கலாம்: ஃப்ரீ ரேடிக்கல்கள் மற்றும் நான்ராடிகல்ஸ். ஒன்று அல்லது அதற்கு மேற்பட்ட இணைக்கப்படாத எலக்ட்ரான்களைக் கொண்ட மூலக்கூறுகள் மற்றும் மூலக்கூறுக்கு வினைத்திறனைக் கொடுப்பது ஃப்ரீ ரேடிக்கல்கள் என்று அழைக்கப்படுகின்றன. 2 ஃப்ரீ ரேடிக்கல்கள் அவற்றின் இணைக்கப்படாத எலக்ட்ரான்களைப் பகிர்ந்து கொள்ளும்போது, தீவிரமற்ற வடிவங்கள் உருவாக்கப்படுகின்றன. உடலியல் முக்கியத்துவம் வாய்ந்த 3 முக்கிய ROS ஆனது சூப்பர் ஆக்சைடு அனான் (O22.), ஹைட்ராக்சில் ரேடிக்கல் (OH) மற்றும் ஹைட்ரஜன் பெராக்சைடு (H2O2) ஆகும். ROS அட்டவணை 1 இல் சுருக்கப்பட்டுள்ளது.
மூலக்கூறு ஆக்ஸிஜனுடன் 1 எலக்ட்ரானைச் சேர்ப்பதன் மூலம் சூப்பர் ஆக்சைடு அயனி உருவாகிறது.22 இந்த செயல்முறை நிகோடின் அடினைன் டைனுக்ளியோடைடு பாஸ்பேட் [NAD(P)H] ஆக்சிடேஸ் அல்லது சாந்தைன் ஆக்சிடேஸ் அல்லது மைட்டோகாண்ட்ரியல் எலக்ட்ரான் டிரான்ஸ்போர்ட் சிஸ்டம் மூலம் மத்தியஸ்தம் செய்யப்படுகிறது. சூப்பர் ஆக்சைடு அயனியை உற்பத்தி செய்வதற்கான முக்கிய தளம் மைட்டோகாண்ட்ரியா ஆகும், இது அடினோசின் ட்ரைபாஸ்பேட்டை உற்பத்தி செய்வதற்கான கலத்தின் இயந்திரமாகும். பொதுவாக, எலக்ட்ரான்கள் மைட்டோகாண்ட்ரியல் எலக்ட்ரான் போக்குவரத்து சங்கிலி மூலம் ஆக்ஸிஜனைக் குறைப்பதற்காக மாற்றப்படுகின்றன, ஆனால் தோராயமாக 1 முதல் 3% எலக்ட்ரான்கள் கணினியிலிருந்து கசிந்து சூப்பர் ஆக்சைடை உருவாக்குகின்றன. NAD(P)H ஆக்சிடேஸ் பாலிமார்போநியூக்ளியர் லுகோசைட்டுகள், மோனோசைட்டுகள் மற்றும் மேக்ரோபேஜ்களில் காணப்படுகிறது. பாகோசைட்டோசிஸில், இந்த செல்கள் சூப்பர் ஆக்சைடு வெடிப்பை உருவாக்குகின்றன, இது பாக்டீரிசைடு செயல்பாட்டிற்கு வழிவகுக்கிறது. சூப்பர் ஆக்சைடு டிஸ்முடேஸ் (SODs, EC 1.15.1.1) செயல்பாட்டின் மூலம் ஹைட்ரஜன் பெராக்சைடாக மாற்றப்படுகிறது. ஹைட்ரஜன் பெராக்சைடு பிளாஸ்மா சவ்வு முழுவதும் எளிதில் பரவுகிறது. ஹைட்ரஜன் பெராக்சைடு சாந்தைன் ஆக்சிடேஸ், அமினோ அமில ஆக்சிடேஸ், மற்றும் NAD(P)H ஆக்சிடேஸ்23,24 மற்றும் பெராக்ஸிசோம்களில் மூலக்கூறு ஆக்ஸிஜனை வளர்சிதை மாற்ற எதிர்வினைகளில் உற்பத்தி செய்கிறது. ஹேபர்-வெயிஸ் மற்றும் ஃபென்டன் வினைகள் எனப்படும் வினைகளின் தொடர்ச்சியாக, Fe2 அல்லது Cu2 போன்ற பரிமாற்ற உலோகங்களின் முன்னிலையில் H2O21 OH21.25 ஆக உடைந்துவிடும்.
Fe31 +¸.O2′?Fe2 +′O2 ஹேபர் வெயிஸ்
Fe2 +′H2O2′?Fe3 +′OH′+ .OH ஃபென்டன் எதிர்வினை
O 2 தானே H2 O2 உடன் வினைபுரிந்து OH' ஐ உருவாக்குகிறது.26,27 ஹைட்ராக்சில் ரேடிக்கல் ROS இன் மிகவும் வினைத்திறன் கொண்டது மற்றும் புரதங்கள், கொழுப்புகள் மற்றும் கார்போஹைட்ரேட்டுகள் மற்றும் DNA ஐ சேதப்படுத்தும். பாலிஅன்சாச்சுரேட்டட் கொழுப்பு அமிலங்களிலிருந்து எலக்ட்ரானை எடுத்துக்கொள்வதன் மூலமும் இது லிப்பிட் பெராக்ஸைடேஷனைத் தொடங்கலாம்.
கிரானுலோசைடிக் என்சைம்கள் ஈசினோபில் பெராக்ஸிடேஸ் மற்றும் மைலோபெராக்ஸிடேஸ் (எம்பிஓ) வழியாக H2O2 இன் வினைத்திறனை மேலும் விரிவுபடுத்துகின்றன. செயல்படுத்தப்பட்ட நியூட்ரோபில்களில், H2O2 MPO ஆல் உட்கொள்ளப்படுகிறது. குளோரைடு அயனியின் முன்னிலையில், H2O2 ஹைபோகுளோரஸ் அமிலமாக (HOCl) மாற்றப்படுகிறது. HOCl அதிக ஆக்ஸிஜனேற்றம் மற்றும் காற்றுப்பாதைகளில் உள்ள நோய்க்கிருமிகளைக் கொல்வதில் முக்கியப் பங்கு வகிக்கிறது. இருப்பினும், HOCl டிஎன்ஏவுடன் வினைபுரிந்து டிஎன்ஏ புரத தொடர்புகளைத் தூண்டலாம் மற்றும் பைரிமிடின் ஆக்சிஜனேற்றப் பொருட்களை உற்பத்தி செய்து டிஎன்ஏ தளங்களில் குளோரைடைச் சேர்க்கலாம்.28 ஈசினோபில் பெராக்ஸிடேஸ் மற்றும் MPO ஆனது ஆக்சிஜனேற்றம், நைட்ரேஷன் மற்றும் புரோட்டீன் குறுக்கு இணைப்புகள் மூலம் டைரோசில் ரேடிக்கல்கள் மூலம் புரதங்களை மாற்றியமைப்பதன் மூலம் ஆக்ஸிஜனேற்ற அழுத்தத்திற்கு பங்களிக்கிறது.29,30-31
பிற ஆக்ஸிஜன்-பெறப்பட்ட ஃப்ரீ ரேடிக்கல்கள் பெராக்சைல் ரேடிக்கல்கள் (ROO$). இந்த தீவிரவாதிகளின் எளிமையான வடிவம் ஹைட்ரோபெராக்சைல் ரேடிக்கல் (HOO$) மற்றும் கொழுப்பு அமில பெராக்ஸைடேஷனில் பங்கு வகிக்கிறது. ஃப்ரீ ரேடிக்கல்கள் பக்கச் சங்கிலி மெத்திலீன் கார்பனில் இருந்து ஹைட்ரஜன் அணுவைச் சுருக்கி லிப்பிட் பெராக்ஸைடேஷன் சங்கிலி எதிர்வினைகளைத் தூண்டலாம். லிப்பிட் ரேடிக்கல் பின்னர் ஆக்ஸிஜனுடன் வினைபுரிந்து பெராக்சைல் ரேடிக்கலை உருவாக்குகிறது. பெராக்சைல் ரேடிக்கல் ஒரு சங்கிலி எதிர்வினையைத் தொடங்குகிறது மற்றும் பாலிஅன்சாச்சுரேட்டட் கொழுப்பு அமிலங்களை லிப்பிட் ஹைட்ரோபெராக்சைடுகளாக மாற்றுகிறது. லிப்பிட் ஹைட்ரோபெராக்சைடுகள் மிகவும் நிலையற்றவை மற்றும் ஆல்டிஹைடுகள் (4-ஹைட்ராக்ஸி-2,3-நோனெனல் போன்றவை) மற்றும் மலோண்டியால்டிஹைடுகள் (எம்டிஏக்கள்) போன்ற இரண்டாம் நிலை தயாரிப்புகளாக எளிதில் சிதைந்துவிடும். ஐசோப்ரோஸ்டேன்ஸ் என்பது லிப்பிட் பெராக்சிடேஷன் தயாரிப்புகளின் மற்றொரு குழுவாகும், அவை அராச்சிடோனிக் அமிலத்தின் பெராக்ஸைடேஷன் மூலம் உருவாக்கப்படுகின்றன, மேலும் ஆஸ்துமாவின் பிளாஸ்மா மற்றும் மூச்சுத் திணறல்களில் உயர்த்தப்படுவது கண்டறியப்பட்டது. சவ்வு அமைப்பு.
ஹைட்ரஜன் பெராக்சைடு, சூப்பர் ஆக்சைடு ரேடிக்கல், ஆக்சிஜனேற்றப்பட்ட குளுதாதயோன் (ஜிஎஸ்எஸ்ஜி), எம்டிஏக்கள், ஐசோப்ரோஸ்டேன்ஸ்கள், கார்போனைல்கள் மற்றும் நைட்ரோடைரோசின் ஆகியவை பிளாஸ்மா, இரத்தம் அல்லது மூச்சுக்குழாய் அழற்சி மாதிரிகளிலிருந்து ஆக்சிஜனேற்றத்தின் பயோமார்க்ஸர்களாக தரநிலைப்படுத்தப்பட்ட மதிப்பீடுகளால் எளிதாக அளவிட முடியும்.
ஆக்ஸிஜனேற்றத்தின் வெளிப்புற ஆதாரம்
சிகரெட் புகை
சிகரெட் புகையில் பல ஆக்ஸிஜனேற்றங்கள் மற்றும் ஃப்ரீ ரேடிக்கல்கள் மற்றும் சூப்பர் ஆக்சைடு மற்றும் நைட்ரிக் ஆக்சைடு போன்ற கரிம சேர்மங்கள் உள்ளன. மேலும், சிகரெட் புகையை நுரையீரலில் உள்ளிழுப்பது நியூட்ரோபில்கள் மற்றும் மேக்ரோபேஜ்கள் போன்ற சில எண்டோஜெனஸ் வழிமுறைகளை செயல்படுத்துகிறது, இது ஆக்ஸிஜனேற்ற காயத்தை மேலும் அதிகரிக்கிறது. .
ஓசோன் வெளிப்பாடு
ஓசோன் வெளிப்பாடு லிப்பிட் பெராக்சிடேஷன் மற்றும் நியூட்ரோபில்களின் ஊடுருவலை காற்றுப்பாதை எபிட்டிலியத்தில் தூண்டலாம். ஓசோனின் குறுகிய கால வெளிப்பாடு MPO, ஈசினோபில் கேஷனிக் புரதங்கள் மற்றும் லாக்டேட் டீஹைட்ரோஜினேஸ் மற்றும் அல்புமின் போன்ற அழற்சி மத்தியஸ்தர்களின் வெளியீட்டையும் ஏற்படுத்துகிறது. துகள்கள் (காற்றில் இடைநிறுத்தப்பட்ட திட துகள்கள் மற்றும் திரவத் துளிகளின் கலவை) ஆக்ஸிஜனைக் குறைப்பதை ஊக்குவிக்கிறது.
ஹைபோரெக்ஸியா
ஹைபராக்ஸியா என்பது நுரையீரல் அல்லது பிற உடல் திசுக்களில் உள்ள ஆக்ஸிஜனின் இயல்பான பகுதி அழுத்தத்தை விட அதிக ஆக்ஸிஜன் அளவுகளின் நிலைகளைக் குறிக்கிறது. இது எதிர்வினை ஆக்ஸிஜன் மற்றும் நைட்ரஜன் இனங்களின் அதிக உற்பத்திக்கு வழிவகுக்கிறது.40,41
அயனியாக்கும் கதிர்வீச்சு
அயனியாக்கும் கதிர்வீச்சு, O2 முன்னிலையில், ஹைட்ராக்சில் ரேடிக்கல், சூப்பர் ஆக்சைடு மற்றும் ஆர்கானிக் ரேடிக்கல்களை ஹைட்ரஜன் பெராக்சைடு மற்றும் ஆர்கானிக் ஹைட்ரோபெராக்சைடுகளாக மாற்றுகிறது. இந்த ஹைட்ரோபெராக்சைடு இனங்கள் ஃபென்டன் எதிர்வினைகள் வழியாக Fe மற்றும் Cu போன்ற ரெடாக்ஸ் செயலில் உள்ள உலோக அயனிகளுடன் வினைபுரிந்து ஆக்ஸிஜனேற்ற அழுத்தத்தைத் தூண்டுகின்றன. 42,43 நாராயணன் மற்றும் பலர் ஆல்பா துகள்களுக்கு வெளிப்படும் ஃபைப்ரோபிளாஸ்ட்கள் உள்செல்லுலார் O44 2. மற்றும் H2O2 இல் குறிப்பிடத்தக்க அதிகரிப்புகளைக் கொண்டிருந்தன. பிளாஸ்மா சவ்வு-பிணைக்கப்பட்ட NADPH ஆக்சிடேஸ் மூலம் உற்பத்தி. ஆக்டிவேட்டர் புரதம்-2 (AP-44), அணுக்கரு காரணி-kB (NF-kB) மற்றும் p1 ஆகியவை செயல்படுத்தப்படுகின்றன, இதன் விளைவாக கதிர்வீச்சு எதிர்வினை தொடர்பான மரபணுக்கள் வெளிப்படுகின்றன. 2-1 புற ஊதா A (UVA) ஃபோட்டான்கள் ஆக்ஸிஜனேற்ற எதிர்வினைகளைத் தூண்டுகின்றன. போர்பிரின்கள், NADPH ஆக்சிடேஸ் மற்றும் ரைபோஃப்ளேவின்கள் போன்ற எண்டோஜெனஸ் ஃபோட்டோசென்சிடைசர்களின் தூண்டுதலால். 2-Oxo-38- dihydroguanine (1-oxoGua) என்பது OH ரேடிக்கல், 1-எலக்ட்ரான் ஆக்சிடென்ட்கள் மற்றும் சிங்கிள்ட் ஆக்சிஜனின் ஆக்சிஜனேற்றத்தால் உருவான முக்கிய UVA-மத்தியஸ்த DNA ஆக்சிஜனேற்றத் தயாரிப்பு ஆகும். இது முக்கியமாக குவானைனுடன் வினைபுரிகிறது. அயனியாக்கும் கதிர்வீச்சின் நேரடி விளைவு மூலம் தனிமைப்படுத்தப்பட்ட டிஎன்ஏவில் உள்ள தீவிர கேஷன் திறமையாக நிகழும் என நிரூபிக்கப்பட்டுள்ளது.53 அயனியாக்கும் கதிர்வீச்சின் வெளிப்பாட்டிற்குப் பிறகு, குளுதாதயோனின் (ஜிஎஸ்ஹெச்) இன் இன்ட்ராசெல்லுலார் அளவு குறுகிய காலத்திற்கு குறைகிறது ஆனால் மீண்டும் அதிகரிக்கிறது.45
கன உலோக அயனிகள்
இரும்பு, தாமிரம், காட்மியம், பாதரசம், நிக்கல், ஈயம் மற்றும் ஆர்சனிக் போன்ற கன உலோக அயனிகள், வினைத்திறன் தீவிரவாதிகளின் உருவாக்கத்தை தூண்டலாம் மற்றும் லிப்பிட் பெராக்சிடேஷன் மற்றும் அணு புரதங்கள் மற்றும் DNA உடன் எதிர்வினை மூலம் நொதி செயல்பாடுகளை குறைப்பதன் மூலம் செல்லுலார் சேதத்தை ஏற்படுத்தும்.
உலோக-மத்தியஸ்த ஃப்ரீ ரேடிக்கல் உருவாக்கத்தின் மிக முக்கியமான வழிமுறைகளில் ஒன்று ஃபென்டன்-வகை எதிர்வினை வழியாகும். சூப்பர் ஆக்சைடு அயனி மற்றும் ஹைட்ரஜன் பெராக்சைடு இரும்பு மற்றும் தாமிரம் போன்ற மாறுதல் உலோகங்களுடன் தொடர்பு கொள்ளலாம், உலோக வினையூக்கிய ஹேபர்-வெயிஸ்/ஃபென்டன் எதிர்வினை மூலம் OH ரேடிக்கல்களை உருவாக்குகிறது.
ஃபென்டன்-வகை மற்றும் ஹேபர்-வெயிஸ்-வகை பொறிமுறைகளைத் தவிர, சில உலோக அயனிகள் செல்லுலார் மூலக்கூறுகளுடன் நேரடியாக வினைபுரிந்து தியோல் தீவிரவாதிகள் போன்ற ஃப்ரீ ரேடிக்கல்களை உருவாக்கலாம் அல்லது செல் சிக்னலிங் பாதைகளைத் தூண்டலாம். இந்த தீவிரவாதிகள் மற்ற தியோல் மூலக்கூறுகளுடன் வினைபுரிந்து O22.. O22 ஐ உருவாக்கலாம். H2O2 ஆக மாற்றப்படுகிறது, இது கூடுதல் ஆக்ஸிஜன் தீவிர உற்பத்தியை ஏற்படுத்துகிறது. ஆர்சனைட் போன்ற சில உலோகங்கள், உயிரணுக்களில் தீவிர உற்பத்தி முறைகளை செயல்படுத்துவதன் மூலம் மறைமுகமாக ROS உருவாக்கத்தை தூண்டுகின்றன.56
ஆர்சனிக் என்பது மிகவும் நச்சுத் தனிமம் ஆகும், இதில் சூப்பர் ஆக்சைடு (O2 2), சிங்கிள்ட் ஆக்சிஜன் (1O2), பெராக்சைல் ரேடிக்கல் (ROO), நைட்ரிக் ஆக்சைடு (NO), ஹைட்ரஜன் பெராக்சைடு (H2O2) மற்றும் டைமெதிலார்சினிக் பெராக்சைல் ரேடிக்கல்கள் [( CH3)2AsOO ].57–59 ஆர்சனிக் (III) சேர்மங்கள் ஆக்ஸிஜனேற்ற நொதிகளைத் தடுக்கலாம், குறிப்பாக GSH-சார்ந்த நொதிகளான குளுதாதயோன்-எஸ்-டிரான்ஸ்ஃபெரேஸ்கள் (GSTகள்), குளுதாதயோன் பெராக்ஸிடேஸ் (GSH-Px) மற்றும் GSH ரிடக்டேஸ், பிணைப்பு வழியாக - அவர்களின் சல்பைட்ரைல் (ŠSH) குழுக்களுக்கு.60,61
ஈயம் லிப்பிட் பெராக்சிடேஷனை அதிகரிக்கிறது.62 ஈயம் வெளிப்பட்ட பிறகு திசு SOD மற்றும் மூளை GPx செயல்பாட்டில் குறிப்பிடத்தக்க குறைவுகள் பதிவாகியுள்ளன. ஈய வெளிப்பாடு திசு தியோல்களைப் பாதிப்பதன் மூலம் ஜிஎஸ்டியைத் தடுக்கலாம்.
உலோக-வினையூக்கிய எதிர்வினைகளால் உருவாக்கப்பட்ட ROS டிஎன்ஏ தளங்களை மாற்றியமைக்க முடியும். Fe21, Cu21 மற்றும் Ni21 போன்ற உலோக அயனிகளால் ஆக்ஸிஜனேற்ற சேதத்தின் விளைவாக G / C, G / T மற்றும் C / T ஆகிய மூன்று அடிப்படை மாற்றீடுகள் ஏற்படலாம். Reid et al65, G/C ஆனது பெரும்பாலும் Fe21 ஆல் தயாரிக்கப்பட்டதாகவும், C/T மாற்றீடு Cu21 மற்றும் Ni21 ஆல் தயாரிக்கப்பட்டதாகவும் காட்டியது.
ஆன்டிஆக்ஸிடன்ட்கள்
மனித உடலில் பலவிதமான ஆக்ஸிஜனேற்றங்கள் உள்ளன, அவை ஆக்ஸிஜனேற்றிகளின் விளைவை சமநிலைப்படுத்த உதவுகின்றன. அனைத்து நடைமுறை நோக்கங்களுக்காக, இவற்றை 2 வகைகளாகப் பிரிக்கலாம்: நொதி (அட்டவணை 2) மற்றும் நொதி அல்லாத (அட்டவணை 3).
நொதி ஆக்ஸிஜனேற்றிகள்
நுரையீரலின் முக்கிய நொதி ஆக்ஸிஜனேற்றிகள் SODகள் (EC 1.15.1.11), கேடலேஸ் (EC 1.11.1.6) மற்றும் GSH-Px (EC 1.11.1.9) ஆகும். இந்த முக்கிய நொதிகளுக்கு மேலதிகமாக, ஹீம் ஆக்சிஜனேஸ்-1 (EC 1.14.99.3) மற்றும் ரெடாக்ஸ் புரதங்கள், தியோரெடாக்சின்கள் (TRXs, EC 1.8.4.10), பெராக்சிரெடாக்சின்கள் (PRXs, EC 1.11.1.15) மற்றும் குளுடரெடாக்ஸின்கள் உட்பட பிற ஆக்ஸிஜனேற்றிகள். , நுரையீரல் ஆக்ஸிஜனேற்ற பாதுகாப்புகளில் முக்கிய பங்கு வகிப்பதாகவும் கண்டறியப்பட்டுள்ளது.
சூப்பர் ஆக்சைடு பல்வேறு மூலங்களிலிருந்து உற்பத்தி செய்யப்படும் முதன்மை ROS என்பதால், SOD மூலம் அதன் சிதைவு ஒவ்வொரு கலத்திற்கும் முதன்மை முக்கியத்துவம் வாய்ந்தது. SOD இன் அனைத்து 3 வடிவங்களும், அதாவது CuZn-SOD, Mn-SOD மற்றும் EC-SOD ஆகியவை மனித நுரையீரலில் பரவலாக வெளிப்படுத்தப்படுகின்றன. Mn-SOD மைட்டோகாண்ட்ரியா மேட்ரிக்ஸில் உள்ளமைக்கப்படுகிறது. EC-SOD முதன்மையாக எக்ஸ்ட்ராசெல்லுலர் மேட்ரிக்ஸில் உள்ளமைக்கப்படுகிறது, குறிப்பாக அதிக அளவு வகை I கொலாஜன் இழைகள் மற்றும் நுரையீரல் மற்றும் சிஸ்டமிக் நாளங்களைச் சுற்றியுள்ள பகுதிகளில். இது மூச்சுக்குழாய் எபிட்டிலியம், அல்வியோலர் எபிட்டிலியம் மற்றும் அல்வியோலர் மேக்ரோபேஜ்களிலும் கண்டறியப்பட்டது. 66,67 ஒட்டுமொத்தமாக, CuZn- SOD மற்றும் Mn-SOD ஆகியவை பொதுவாக சூப்பர் ஆக்சைடு ரேடிக்கல்களின் மொத்தத் துப்புரவுகளாகச் செயல்படுவதாகக் கருதப்படுகிறது. நுரையீரலில் உள்ள ஒப்பீட்டளவில் அதிக EC-SOD நிலை அதன் குறிப்பிட்ட பிணைப்பு எக்ஸ்ட்ராசெல்லுலர் மேட்ரிக்ஸ் கூறுகளுடன் நுரையீரல் மேட்ரிக்ஸ் பாதுகாப்பின் ஒரு அடிப்படை அங்கமாக இருக்கலாம்.68
SODகளின் செயல்பாட்டால் அல்லது சாந்தைன் ஆக்சிடேஸ் போன்ற ஆக்சிடேஸ்களின் செயல்பாட்டால் உற்பத்தி செய்யப்படும் H2O2, கேடலேஸ் மற்றும் GSH-Px மூலம் தண்ணீராகக் குறைக்கப்படுகிறது. கேடலேஸ் 4 ஒரே மாதிரியான மோனோமர்களைக் கொண்ட ஒரு டெட்ராமராக உள்ளது, ஒவ்வொன்றும் செயலில் உள்ள தளத்தில் ஒரு ஹீம் குழுவைக் கொண்டுள்ளது. கேடலேஸ்-ஃபெரிகாடலேஸ் (தண்ணீருடன் ஒருங்கிணைக்கப்பட்ட இரும்பு) மற்றும் கலவை I (ஆக்சிஜன் அணுவுடன் சிக்கலான இரும்பு) ஆகியவற்றின் 2 இணக்கங்களுக்கு இடையில் மாற்றுவதன் மூலம் H2O2 இன் சிதைவு செய்யப்படுகிறது. கேடலேஸ் NADPH ஐ ஒரு குறைக்கும் சமமாக பிணைக்கிறது, அது தண்ணீராக குறைக்கப்படுவதால், H2O2 மூலம் நொதியின் ஆக்ஸிஜனேற்ற செயலிழப்பைத் தடுக்கிறது.69
ரெடாக்ஸ் சுழற்சியில் உள்ள என்சைம்கள் H2O2 மற்றும் லிப்பிட் ஹைட்ரோபெராக்சைடுகள் (மெம்ப்ரேன் லிப்பிட் பெராக்ஸைடேஷனின் விளைவாக உருவாகிறது) ஆகியவற்றின் குறைப்புக்கு காரணமானது, GSH-Pxs.70 GSH-Pxs என்பது டெட்ராமெரிக் என்சைம்களின் குடும்பமாகும். செயலில் உள்ள தளங்கள் மற்றும் H2O2 மற்றும் லிப்பிட் பெராக்சைடுகளை அவற்றின் தொடர்புடைய ஆல்கஹால்களுக்கு குறைக்க GSH போன்ற குறைந்த-மூலக்கூறு-எடை தியோல்களைப் பயன்படுத்துகின்றன. நான்கு GSH- Pxs விவரிக்கப்பட்டுள்ளன, வெவ்வேறு மரபணுக்களால் குறியிடப்பட்டது: GSH- Px-1 (செல்லுலார் GSH-Px) எங்கும் உள்ளது மற்றும் H2O2 மற்றும் கொழுப்பு அமில பெராக்சைடுகளைக் குறைக்கிறது, ஆனால் பெராக்சைல் லிப்பிட்கள் அல்ல. -Px-71 (பாஸ்போலிப்பிட் ஹைட்ரோபெராக்சைடு GSH-Px), இது பல்வேறு குறைந்த-மூலக்கூறு-எடை தியோல்களைக் குறைப்பதற்கு சமமானதாகப் பயன்படுத்தலாம். GSH-Px-4 (இரைப்பை குடல் GSH-Px) இரைப்பை குடல் எபிடெலியல் செல்களில் உள்ளமைக்கப்படுகிறது, அங்கு உணவு பெராக்சைடுகளைக் குறைக்க உதவுகிறது. எக்ஸ்ட்ராசெல்லுலர் பெட்டி மற்றும் பாலூட்டிகளில் மிக முக்கியமான எக்ஸ்ட்ராசெல்லுலர் ஆக்ஸிஜனேற்ற நொதிகளில் ஒன்றாக நம்பப்படுகிறது. இவற்றில், எக்ஸ்ட்ராசெல்லுலர் GSH-Px மனித நுரையீரலில் மிகவும் பரவலாக ஆராயப்படுகிறது.2
கூடுதலாக, H2O2 ஐ அகற்றுவது பல தியோல்-கொண்ட என்சைம்களுடன் நெருக்கமாக தொடர்புடையது, அதாவது, TRXs (TRX1 மற்றும் TRX2), தியோரெடாக்சின் ரிடக்டேஸ்கள் (EC 1.8.1.9) (TRRகள்), PRXs (தியோரெடாக்சின் பெராக்சிடேஸ்கள்) மற்றும் குளுடரிடாக்ஸின்கள்.74.
இரண்டு டிஆர்எக்ஸ் மற்றும் டிஆர்ஆர்கள் மனித உயிரணுக்களில் வகைப்படுத்தப்பட்டுள்ளன, அவை சைட்டோசோல் மற்றும் மைட்டோகாண்ட்ரியா இரண்டிலும் உள்ளன. நுரையீரலில், TRX மற்றும் TRR ஆகியவை மூச்சுக்குழாய் மற்றும் அல்வியோலர் எபிட்டிலியம் மற்றும் மேக்ரோபேஜ்களில் வெளிப்படுத்தப்படுகின்றன. மனித உயிரணுக்களில் ஆறு வெவ்வேறு PRXகள் கண்டறியப்பட்டுள்ளன, அவற்றின் அல்ட்ராஸ்ட்ரக்சுரல் கம்பார்ட்மென்டலைசேஷனில் வேறுபடுகின்றன. அல்வியோலர் எபிட்டிலியத்தின் பாதுகாப்பில் PRX VI இன் முக்கியத்துவத்தை பரிசோதனை ஆய்வுகள் வெளிப்படுத்தியுள்ளன. மனித நுரையீரல் அனைத்து PRX களையும் மூச்சுக்குழாய் எபிட்டிலியம், அல்வியோலர் எபிட்டிலியம் மற்றும் மேக்ரோபேஜ்களில் வெளிப்படுத்துகிறது. 75 PRX V சமீபத்தில் ஒரு பெராக்ஸைனிட்ரைட் ரிடக்டேஸாக செயல்படுவதாகக் கண்டறியப்பட்டது .76
இந்த ஆன்டிஆக்ஸிடன்ட்களுக்கு பொதுவானது NADPH இன் தேவை குறைக்கும் சமமானதாகும். NADPH செயலில் உள்ள வடிவத்தில் வினையூக்கியை பராமரிக்கிறது மற்றும் TRX மற்றும் GSH ரிடக்டேஸ் (EC 1.6.4.2) மூலம் ஒரு இணைப்பாளராகப் பயன்படுத்தப்படுகிறது, இது GSSG ஐ GSH ஆக மாற்றுகிறது, இது GSH-Pxsக்கான இணை மூலக்கூறு ஆகும். குளுக்கோஸ்-1-பாஸ்பேட்டை 6-பாஸ்போகுளோகோனோலாக்டோனாக மாற்றும் போது, பென்டோஸ் பாஸ்பேட் பாதையின் முதல் மற்றும் விகிதத்தைக் கட்டுப்படுத்தும் நொதியான குளுக்கோஸ்-6-பாஸ்பேட் டீஹைட்ரோஜினேஸ் மூலம் NADP6 ஐக் குறைப்பதன் மூலம் உள்செல்லுலார் NADPH உருவாக்கப்படுகிறது. NADPH ஐ உருவாக்குவதன் மூலம், குளுக்கோஸ்-6-பாஸ்பேட் டீஹைட்ரஜனேஸ் என்பது சைட்டோசோலிக் ஜிஎஸ்ஹெச் இடையகத் திறனை (ஜிஎஸ்ஹெச்/ஜிஎஸ்எஸ்ஜி) ஒரு முக்கியமான நிர்ணயிப்பதாகும், எனவே, ஒரு அத்தியாவசிய, ஒழுங்குமுறை ஆக்ஸிஜனேற்ற நொதியாகக் கருதலாம்.78,79
GSTகள் (EC 2.5.1.18), மற்றொரு ஆக்ஸிஜனேற்ற நொதி குடும்பம், நிறைவுறா ஆல்டிஹைடுகள், எபோக்சைடுகள் மற்றும் ஹைட்ரோபெராக்சைடுகள் போன்ற இரண்டாம் நிலை வளர்சிதை மாற்றங்களை செயலிழக்கச் செய்கிறது. ஜிஎஸ்டியின் மூன்று முக்கிய குடும்பங்கள் விவரிக்கப்பட்டுள்ளன: சைட்டோசோலிக் ஜிஎஸ்டி, மைட்டோகாண்ட்ரியல் ஜிஎஸ்டி,80,81 மற்றும் சவ்வு-தொடர்புடைய மைக்ரோசோமல் ஜிஎஸ்டி ஆகியவை ஈகோசனாய்டு மற்றும் ஜிஎஸ்ஹெச் வளர்சிதை மாற்றத்தில் பங்கு வகிக்கின்றன. Pi, Sigma, Theta, Omega, and Zeta.82–83 அழுத்தமில்லாத நிலைகளின் போது, வகுப்பு Mu மற்றும் Pi GSTகள் முறையே Ask86 மற்றும் JNK ஆகிய கைனேஸ்களுடன் தொடர்பு கொள்கின்றன, மேலும் இந்த கைனேஸ்களைத் தடுக்கின்றன. ஆக்சிஜனேற்ற அழுத்தத்திற்கு பதில் JNK.
நொன்சைமடிக் ஆன்டிஆக்ஸிடன்ட்கள்
நொன்சைமடிக் ஆன்டிஆக்ஸிடன்ட்களில் வைட்டமின்கள் (வைட்டமின்கள் சி மற்றும் ஈ), பி-கரோட்டின், யூரிக் அமிலம் மற்றும் ஜிஎஸ்ஹெச், டிரிபெப்டைட் (எல்ஜி-குளூட்டமைல்-எல்-சிஸ்டைனைல்-எல்-கிளைசின்) போன்ற குறைந்த-மூலக்கூறு-எடை கலவைகள் அடங்கும். சல்பைட்ரைல்) குழு.
வைட்டமின் சி (அஸ்கார்பிக் அமிலம்)
நீரில் கரையக்கூடிய வைட்டமின் சி (அஸ்கார்பிக் அமிலம்) முதன்மையாக ஆக்சிஜன் ஃப்ரீ ரேடிக்கல்களை அகற்றுவதன் மூலம் உள்செல்லுலார் மற்றும் எக்ஸ்ட்ராசெல்லுலர் அக்வஸ்-ஃபேஸ் ஆக்ஸிஜனேற்ற திறனை வழங்குகிறது. இது வைட்டமின் ஈ ஃப்ரீ ரேடிக்கல்களை மீண்டும் வைட்டமின் ஈ ஆக மாற்றுகிறது. அதன் பிளாஸ்மா அளவுகள் வயதுக்கு ஏற்ப குறைவதாகக் காட்டப்பட்டுள்ளது.91,92
வைட்டமின் ஈ (ஏ-டோகோபெரோல்)
லிப்பிட்-கரையக்கூடிய வைட்டமின் ஈ செல் சவ்வின் ஹைட்ரோபோபிக் உட்புறத்தில் செறிவூட்டப்பட்டுள்ளது மற்றும் ஆக்ஸிஜனேற்றத்தால் தூண்டப்பட்ட சவ்வு காயத்திற்கு எதிரான முக்கிய பாதுகாப்பு ஆகும். வைட்டமின் ஈ பெராக்சில் ரேடிக்கலுக்கு எலக்ட்ரானை தானம் செய்கிறது, இது லிப்பிட் பெராக்ஸைடேஷனின் போது உற்பத்தி செய்யப்படுகிறது. a-டோகோபெரோல் என்பது வைட்டமின் E இன் மிகவும் செயலில் உள்ள வடிவம் மற்றும் உயிரணுவில் உள்ள முக்கிய சவ்வு-பிணைப்பு ஆக்ஸிஜனேற்றமாகும். வைட்டமின் ஈ புற்றுநோய் செல்களின் அப்போப்டொசிஸைத் தூண்டுகிறது மற்றும் ஃப்ரீ ரேடிக்கல் அமைப்புகளைத் தடுக்கிறது.93
குளுதாதயோன்
அனைத்து செல் பெட்டிகளிலும் GSH அதிகமாக உள்ளது மற்றும் கரையக்கூடிய முக்கிய ஆக்ஸிஜனேற்றியாகும். GSH/GSSG விகிதம் ஆக்ஸிஜனேற்ற அழுத்தத்தின் முக்கிய நிர்ணயம் ஆகும். GSH பல வழிகளில் அதன் ஆக்ஸிஜனேற்ற விளைவுகளைக் காட்டுகிறது. GSH அதன் எலக்ட்ரானை H94O2 க்கு தானமாக H2O மற்றும் O2 ஆக குறைக்கிறது. GSSG மீண்டும் GSH ரிடக்டேஸால் GSH ஆக குறைக்கப்படுகிறது, இது NAD(P)Hஐ எலக்ட்ரான் தானமாகப் பயன்படுத்துகிறது. லிப்பிட் பெராக்சிடேஷனில் இருந்து உயிரணு சவ்வுகளைப் பாதுகாப்பதற்கும் GSH-Pxs முக்கியமானவை. குறைக்கப்பட்ட குளுதாதயோன் சவ்வு லிப்பிடுகளுக்கு புரோட்டான்களை நன்கொடையாக அளித்து ஆக்ஸிஜனேற்ற தாக்குதல்களிலிருந்து பாதுகாக்கிறது.2
GSH என்பது GSH-Px மற்றும் Transferase போன்ற பல நச்சு நீக்கும் நொதிகளுக்கு ஒரு இணை காரணியாகும். வைட்டமின் சி மற்றும் ஈ ஆகியவற்றை அவற்றின் செயலில் உள்ள வடிவங்களுக்கு மாற்றுவதில் இது ஒரு பங்கு வகிக்கிறது. ப்ரோபோப்டோடிக் மற்றும் ஆன்டிபாப்டோடிக் சிக்னலிங் பாதைகளுடன் தொடர்புகொள்வதன் மூலம் அப்போப்டொசிஸுக்கு எதிராக செல்களை GSH பாதுகாக்கிறது.
கரோட்டினாய்டுகள் (பி-கரோட்டின்)
கரோட்டினாய்டுகள் தாவரங்களில் காணப்படும் நிறமிகள். முதன்மையாக, பி-கரோட்டின் பெராக்சில் (ROO), ஹைட்ராக்சில் (OH), மற்றும் சூப்பர் ஆக்சைடு (O22.) ரேடிக்கல்களுடன் வினைபுரிவதாகக் கண்டறியப்பட்டுள்ளது. செறிவுகள்.96 கரோட்டினாய்டுகள் மற்றும் ரெட்டினோயிக் அமிலங்கள் (RAs) இரண்டும் டிரான்ஸ்கிரிப்ஷன் காரணிகளைக் கட்டுப்படுத்தும் திறன் கொண்டவை.97 b-கரோட்டின் ஆக்சிடன்ட்-தூண்டப்பட்ட NF-kB செயல்படுத்தலைத் தடுக்கிறது மற்றும் இன்டர்லூகின் (IL)-98 மற்றும் கட்டி நசிவு காரணி-a உற்பத்தியைத் தடுக்கிறது. கரோட்டினாய்டுகள் செல்களின் அப்போப்டொசிஸையும் பாதிக்கிறது. RA இன் ஆன்டிபிரோலிஃபெரேடிவ் விளைவுகள் பல ஆய்வுகளில் காட்டப்பட்டுள்ளன. RA இன் இந்த விளைவு முக்கியமாக ரெட்டினோயிக் அமில ஏற்பிகளால் மத்தியஸ்தம் செய்யப்படுகிறது மற்றும் செல் வகைகளில் வேறுபடுகிறது. மார்பக புற்றுநோய் உயிரணுக்களில், ரெட்டினோயிக் அமிலம் ஏற்பியானது செல் சுழற்சி தடுப்பு, அப்போப்டொசிஸ் அல்லது இரண்டையும் தூண்டுவதன் மூலம் வளர்ச்சித் தடுப்பைத் தூண்டுவதாகக் காட்டப்பட்டது.6
ஆக்ஸிஜனேற்ற அழுத்தத்தின் விளைவு: மரபணு, உடலியல் மற்றும் உயிர்வேதியியல் இயக்கவியல்
ஆக்ஸிஜனேற்றிகள் மற்றும் ROS க்கு இடையேயான சமநிலை பாதிக்கப்படும் போது ஆக்ஸிஜனேற்ற அழுத்தம் ஏற்படுகிறது, ஏனெனில் ஆன்டிஆக்ஸிடன்ட்கள் குறைதல் அல்லது ROS திரட்சி. ஆக்ஸிஜனேற்ற அழுத்தம் ஏற்படும் போது, உயிரணுக்கள் ஆக்ஸிஜனேற்ற விளைவுகளை எதிர்க்க முயல்கின்றன மற்றும் தற்காப்பு நொதிகள், டிரான்ஸ்கிரிப்ஷன் காரணிகள் மற்றும் கட்டமைப்பு புரதங்களின் மரபணுக்களை செயல்படுத்துதல் அல்லது அமைதிப்படுத்துவதன் மூலம் ரெடாக்ஸ் சமநிலையை மீட்டெடுக்கின்றன. உடலில் ஆக்சிஜனேற்ற அழுத்தத்தை தீர்மானிக்கும் முக்கிய காரணிகள். உடலில் ROS இன் அதிக உற்பத்தி DNA கட்டமைப்பை மாற்றலாம், இதன் விளைவாக புரதங்கள் மற்றும் லிப்பிட்களின் மாற்றம், பல அழுத்தத்தால் தூண்டப்பட்ட டிரான்ஸ்கிரிப்ஷன் காரணிகளை செயல்படுத்துதல் மற்றும் அழற்சிக்கு சார்பான மற்றும் அழற்சி எதிர்ப்பு சைட்டோகைன்களின் உற்பத்தி ஆகியவை ஏற்படலாம்.
டிஎன்ஏ மீது ஆக்ஸிஜனேற்ற அழுத்தத்தின் விளைவுகள்
ROS பல வழிகளில் டிஎன்ஏ மாற்றங்களுக்கு வழிவகுக்கும், இதில் அடிப்படைகளின் சிதைவு, ஒற்றை அல்லது இரட்டை இழைகள் கொண்ட டிஎன்ஏ முறிவுகள், பியூரின், பைரிமிடின் அல்லது சர்க்கரை-பிணைப்பு மாற்றங்கள், பிறழ்வுகள், நீக்குதல்கள் அல்லது இடமாற்றங்கள் மற்றும் புரதங்களுடன் குறுக்கு இணைப்பு ஆகியவை அடங்கும். இந்த டிஎன்ஏ மாற்றங்களில் பெரும்பாலானவை (படம் 1) கார்சினோஜெனிசிஸ், வயதான மற்றும் நரம்பியக்கடத்தல், இருதய மற்றும் தன்னுடல் தாக்க நோய்களுக்கு மிகவும் பொருத்தமானவை. புகையிலை புகை, ரெடாக்ஸ் உலோகங்கள் மற்றும் இரும்பு, காட்மியம், குரோம் மற்றும் ஆர்சனிக் போன்ற ரெடாக்ஸ் அல்லாத உலோகங்கள், ஃப்ரீ ரேடிக்கல்களை உருவாக்குவதன் மூலம் அல்லது தியோல் குழுக்களுடன் பிணைப்பதன் மூலம் புற்றுநோய் மற்றும் வயதானதில் ஈடுபட்டுள்ளன. 8-OH-G இன் உருவாக்கம் என்பது ஆக்ஸிஜனேற்ற அழுத்தத்தின் மூலம் நிகழும் சிறந்த அறியப்பட்ட டிஎன்ஏ சேதமாகும், மேலும் இது புற்றுநோயை உண்டாக்குவதற்கான சாத்தியமான உயிரியலாகும்.
மரபணுக்களின் ஊக்குவிப்பு பகுதிகள் டிரான்ஸ்கிரிப்ஷன் காரணிகளுக்கான ஒருமித்த தொடர்களைக் கொண்டிருக்கின்றன. இந்த டிரான்ஸ்கிரிப்ஷன் காரணி-பைண்டிங் தளங்கள் ஆக்சிடன்ட் தாக்குதலுக்கு ஆளாகக்கூடிய ஜிசி நிறைந்த வரிசைகளைக் கொண்டுள்ளன. டிரான்ஸ்கிரிப்ஷன் காரணி பிணைப்பு தளங்களில் 8-OH-G டிஎன்ஏ உருவாக்கம், டிரான்ஸ்கிரிப்ஷன் காரணிகளின் பிணைப்பை மாற்றியமைக்கலாம், இதனால் AP-1 மற்றும் Sp-1 இலக்கு வரிசைகளில் காட்டப்பட்டுள்ளபடி தொடர்புடைய மரபணுக்களின் வெளிப்பாட்டை மாற்றலாம்.103 தவிர 8-OH-G, 8,59 -cyclo-29 -deoxyadenosine (cyclo-dA) TATA பெட்டியில் அமைந்திருந்தால், செல் அமைப்பில் உள்ள ஒரு நிருபர் மரபணுவிலிருந்து படியெடுத்தலைத் தடுப்பதாகவும் காட்டப்பட்டுள்ளது. . TATA-பிணைப்பு புரதத்தின் பிணைப்பு சைக்ளோ-dA இருப்பதால் பலவீனமடையலாம்.
ஆக்ஸிஜனேற்ற அழுத்தமானது மைக்ரோசாட்லைட் (குறுகிய டேன்டெம் ரிபீட்ஸ்) பகுதிகளின் உறுதியற்ற தன்மையை ஏற்படுத்துகிறது. ரெடாக்ஸ் ஆக்டிவ் மெட்டல் அயனிகள், ஹைட்ராக்ஸைல் ரேடிக்கல்கள் மைக்ரோசாட்லைட் உறுதியற்ற தன்மையை அதிகரிக்கின்றன.105 ஆக்சிடன்ட் காயத்தால் ஏற்படும் ஒற்றை இழை டிஎன்ஏ முறிவுகளை செல்கள் எளிதில் பொறுத்துக்கொள்ள முடியும் என்றாலும், அயனியாக்கும் கதிர்வீச்சினால் தூண்டப்படும் இரட்டை இழை டிஎன்ஏ முறிவுகள் உயிரணு உயிர்வாழ்வுக்கு குறிப்பிடத்தக்க அச்சுறுத்தலாக இருக்கலாம்.106
டிஎன்ஏவில் உள்ள சிபிஜி தீவுகளில் மெத்திலேஷன் என்பது ஒரு முக்கியமான எபிஜெனெடிக் பொறிமுறையாகும், இது மரபணு அமைதியை ஏற்படுத்தக்கூடும். 5-MeCyt முதல் 5-ஹைட்ராக்சிமீதில் யுரேசில் (5-OHMeUra) ஆக்சிஜனேற்றம் தைமின் அல்லது 5-ஹைட்ராக்ஸிமெதில் சைட்டோசின் இடைநிலைகளின் டீமினேஷன்/ஆக்சிடேஷன் வினைகள் மூலம் ஏற்படலாம் ஆக்ஸிஜனேற்ற தாக்குதல்களால் தூண்டப்பட்ட பிறழ்ந்த டிஎன்ஏ மெத்திலேஷன் முறைகள் டிஎன்ஏ பழுதுபார்க்கும் செயல்பாட்டையும் பாதிக்கிறது.
லிப்பிட்களில் ஆக்ஸிஜனேற்ற அழுத்தத்தின் விளைவுகள்
ROS லிப்பிட் பெராக்சிடேஷனைத் தூண்டலாம் மற்றும் சவ்வு-பிணைப்பு ஏற்பிகள் மற்றும் என்சைம்களை செயலிழக்கச் செய்யலாம் மற்றும் திசு ஊடுருவலை அதிகரிக்கலாம். -இணைப்புகள்.109-110 112-ஹைட்ராக்ஸி-4-நோனேனல் உள்செல்லுலார் ஜிஎஸ்ஹெச் குறைவை ஏற்படுத்துகிறது மற்றும் பெராக்சைடு உற்பத்தியைத் தூண்டுகிறது, 2 மேல்தோல் வளர்ச்சி காரணி ஏற்பியை செயல்படுத்துகிறது, 113,114 மற்றும் ஃபைப்ரோனெக்டின் உற்பத்தியைத் தூண்டுகிறது. , ஆக்சிஜனேற்ற அழுத்தத்தின் மறைமுக பயோமார்க்ஸர்களாகப் பயன்படுத்தப்படுகின்றன, மேலும் அதிகரித்த அளவுகள் வெளியேற்றப்படும் சுவாசக் கண்டன்சேட் அல்லது மூச்சுக்குழாய் அழற்சி திரவம் அல்லது நாள்பட்ட தடுப்பு நுரையீரல் நோய் நோயாளிகள் அல்லது புகைப்பிடிப்பவர்களின் நுரையீரலில் காட்டப்பட்டது.115-116
புரதங்களில் ஆக்ஸிஜனேற்ற அழுத்தத்தின் விளைவுகள்
ROS ஆனது பெப்டைட் சங்கிலியின் துண்டாடுதல், புரதங்களின் மின் கட்டணத்தை மாற்றுதல், புரதங்களின் குறுக்கு இணைப்பு மற்றும் குறிப்பிட்ட அமினோ அமிலங்களின் ஆக்சிஜனேற்றம் ஆகியவற்றை ஏற்படுத்தலாம். எனவே குறிப்பிட்ட புரோட்டீஸால் சிதைவதன் மூலம் புரோட்டியோலிசிஸுக்கு அதிக பாதிப்புக்கு வழிவகுக்கும். குறிப்பாக ஆக்சிஜனேற்றத்திற்கு ஆக்சிஜனேற்றம் அதிகம். நொதிகளின் ஆக்ஸிஜனேற்ற மாற்றம் அவற்றின் செயல்பாடுகளைத் தடுப்பதாகக் காட்டப்பட்டுள்ளது.120
சில சந்தர்ப்பங்களில், புரதங்களின் குறிப்பிட்ட ஆக்சிஜனேற்றம் நடைபெறலாம். எடுத்துக்காட்டாக, மெத்தியோனைன் மெத்தியோனைன் சல்பாக்சைடு126 ஆகவும் ஃபைனிலாலனைனை ஓ-டைரோசின்127 ஆகவும் ஆக்சிஜனேற்றம் செய்யலாம்; சல்பைட்ரைல் குழுக்கள் ஆக்ஸிஜனேற்றப்பட்டு டைசல்பைட் பிணைப்புகளை உருவாக்கலாம்; 128 மற்றும் கார்போனைல் குழுக்கள் புரதங்களின் பக்க சங்கிலிகளில் அறிமுகப்படுத்தப்படலாம். காமா கதிர்கள், உலோக-வினையூக்கிய ஆக்சிஜனேற்றம், HOCl மற்றும் ஓசோன் ஆகியவை கார்போனைல் குழுக்களை உருவாக்கலாம்.129
சிக்னல் கடத்தலில் ஆக்ஸிஜனேற்ற அழுத்தத்தின் விளைவுகள்
ROS ஆனது சமிக்ஞை கடத்துதலில் ஈடுபட்டுள்ள பல மரபணுக்களின் வெளிப்பாட்டைத் தூண்டலாம். இந்த விகிதத்தின் சீர்குலைவு ரெடாக்ஸ் உணர்திறன் டிரான்ஸ்கிரிப்ஷன் காரணிகளான NF-kB, AP-1,130, செயல்படுத்தப்பட்ட T செல்களின் அணுக்கரு காரணி மற்றும் அழற்சியின் பிரதிபலிப்பில் ஈடுபட்டுள்ள ஹைபோக்ஸியா-தூண்டக்கூடிய காரணி 1 போன்றவற்றை செயல்படுத்துகிறது. ROS வழியாக டிரான்ஸ்கிரிப்ஷன் காரணிகளை செயல்படுத்துவது சிக்னல் டிரான்ஸ்டக்ஷன் கேஸ்கேட்களால் அடையப்படுகிறது, அவை தகவலை வெளியில் இருந்து கலத்தின் உட்புறத்திற்கு அனுப்பும். டைரோசின் கைனேஸ் ஏற்பிகள், எபிடெர்மல் வளர்ச்சி காரணி ஏற்பி, வாஸ்குலர் எண்டோடெலியல் வளர்ச்சி காரணி ஏற்பி, மற்றும் பிளேட்லெட்-பெறப்பட்ட வளர்ச்சி காரணிக்கான ஏற்பி, புரோட்டீன் டைரோசின் பாஸ்பேடேஸ்கள் மற்றும் செரின்/த்ரியோனைன் கைனேஸ்கள் போன்ற வளர்ச்சி காரணி ஏற்பிகள் ROS.1 புற-செல்லுலார் சிக்னல்-ஒழுங்குபடுத்தப்பட்ட கைனேஸ்கள், JNK மற்றும் p131, இவை மைட்டோஜென்-செயல்படுத்தப்பட்ட புரோட்டீன் கைனேஸ் குடும்பத்தின் உறுப்பினர்களாகவும், உயிரணுக்களின் பெருக்கம், வேறுபாடு மற்றும் அப்போப்டொசிஸ் உள்ளிட்ட பல செயல்முறைகளில் ஈடுபட்டுள்ளன, மேலும் அவை ஆக்ஸிஜனேற்றங்களால் கட்டுப்படுத்தப்படலாம்.
ஆக்ஸிஜனேற்ற அழுத்த நிலைமைகளின் கீழ், சி-ஜூனின் டிஎன்ஏ-பிணைப்பு தளத்தில் உள்ள சிஸ்டைன் எச்சங்கள், சில ஏபி-1 துணைக்குழுக்கள் மற்றும் தடுப்பான கேபி கைனேஸ் ஆகியவை மீளக்கூடிய எஸ்-குளுதாதியோலேஷனுக்கு உட்படுகின்றன. NF-kB மற்றும் AP-1, p38 mitogen-activated protein kinase, மற்றும் JNK.134-137 போன்ற ரெடாக்ஸ்-சென்சிட்டிவ் சிக்னலிங் பாதைகளை ஒழுங்குபடுத்துவதில் குளுடரெடாக்சின் மற்றும் TRX முக்கிய பங்கு வகிப்பதாக தெரிவிக்கப்பட்டுள்ளது.
ROS, ஃப்ரீ ரேடிக்கல்கள் மற்றும் UV கதிர்வீச்சு போன்ற ஆக்ஸிஜனேற்ற அழுத்த நிலைமைகளுக்கு பதிலளிக்கும் வகையில் NF-kB செயல்படுத்தப்படலாம். 138 IkB இன் பாஸ்போரைலேஷன் NF-kB ஐ விடுவிக்கிறது மற்றும் மரபணு டிரான்ஸ்கிரிப்ஷனைச் செயல்படுத்த அணுக்கருவிற்குள் நுழைய அனுமதிக்கிறது.139 பல கைனேஸ்கள் உள்ளன. செரின் எச்சங்களில் பாஸ்போரிலேட் IkB களுக்கு தெரிவிக்கப்பட்டது. இந்த கைனேஸ்கள் NF-kB ஐ செயல்படுத்துவதற்கான ஆக்ஸிஜனேற்ற சமிக்ஞைகளின் இலக்குகளாகும். 140 குறைக்கும் முகவர்கள் NF-kB டிஎன்ஏ பிணைப்பை மேம்படுத்துகின்றன, அதேசமயம் ஆக்ஸிஜனேற்ற முகவர்கள் NF-kB இன் DNA பிணைப்பைத் தடுக்கின்றன. NF-kB ஐ ஒழுங்குபடுத்துவதில் TRX 2 எதிர் செயல்களைச் செய்யலாம்: சைட்டோபிளாஸில், இது IkB இன் சிதைவைத் தடுக்கிறது மற்றும் NF-kB செயல்பாட்டைத் தடுக்கிறது, ஆனால் NF-kB டிஎன்ஏ பிணைப்பை நியூக்ளியஸில் மேம்படுத்துகிறது. IkB பல ஆக்ஸிஜனேற்ற பாதுகாப்பு தொடர்பான மரபணுக்களை செயல்படுத்துகிறது. IL-141b, IL-1, கட்டி நெக்ரோசிஸ் காரணி-a, IL-6 மற்றும் பல ஒட்டுதல் மூலக்கூறுகள் போன்ற நோயெதிர்ப்பு மறுமொழியில் பங்கேற்கும் பல மரபணுக்களின் வெளிப்பாட்டை NF-kB ஒழுங்குபடுத்துகிறது. செல்கள் வேறுபாடு.
AP-1 ரெடாக்ஸ் நிலையால் கட்டுப்படுத்தப்படுகிறது. H2O2 முன்னிலையில், சில உலோக அயனிகள் AP-1 ஐ செயல்படுத்த தூண்டலாம். GSH/GSSG இன் விகிதத்தில் அதிகரிப்பு AP-1 பிணைப்பை மேம்படுத்துகிறது, அதே நேரத்தில் GSSG ஆனது AP-1.144 DNA பிணைப்பைத் தடுக்கிறது. புரதங்கள்,145 அதே சமயம் AP-1 இன் DNA பிணைப்பை GSSG மூலம் பல செல் வகைகளில் தடுக்கலாம், சிஸ்டைன் எச்சங்களால் டிசல்பைட் பிணைப்பு உருவாக்கம் AP-1 DNA பிணைப்பைத் தடுக்கிறது.
முடிவுரை
ஆக்சிஜனைப் பயன்படுத்தும் வளர்சிதை மாற்ற எதிர்வினைகளால் ROS இன் அதிகப்படியான உற்பத்தியிலிருந்து ஆக்ஸிஜனேற்ற அழுத்தம் ஏற்படலாம் மற்றும் இடையில் சமநிலையை மாற்றலாம். ஆக்ஸிஜனேற்றம்/ஆக்ஸிஜனேற்ற ஆக்ஸிஜனேற்றங்களுக்கு ஆதரவான நிலைகள். ROS ஆனது செல்லுலார் வளர்சிதை மாற்ற நடவடிக்கைகள் மற்றும் காற்று மாசுபடுத்திகள் அல்லது சிகரெட் புகை போன்ற சுற்றுச்சூழல் காரணிகளால் தயாரிக்கப்படுகிறது. ROS ஆனது, அவற்றின் கட்டமைப்பில் இணைக்கப்படாத எலக்ட்ரான்கள் மற்றும் கார்போஹைட்ரேட்டுகள், நியூக்ளிக் அமிலங்கள், லிப்பிடுகள் மற்றும் புரதங்கள் போன்ற செல்களில் உள்ள பல உயிரியல் மேக்ரோமோலிகுல்களுடன் வினைபுரிந்து அவற்றின் செயல்பாடுகளை மாற்றியமைப்பதால் அதிக வினைத்திறன் கொண்ட மூலக்கூறுகளாகும். ROS ஆனது ரெடாக்ஸ்-சென்சிட்டிவ் டிரான்ஸ்கிரிப்ஷன் காரணிகளை அதிகப்படுத்துவதன் மூலமும், ஹிஸ்டோன் அசிடைலேஷன்/ டீசெடைலேஷன் மாற்றத்தின் மூலம் குரோமாடின் மறுவடிவமைப்பினாலும் பல மரபணுக்களின் வெளிப்பாட்டையும் பாதிக்கிறது. செல் நம்பகத்தன்மை, செயல்படுத்துதல், பெருக்கம் மற்றும் உறுப்பு செயல்பாட்டிற்கு ரெடாக்ஸ் நிலையை ஒழுங்குபடுத்துவது மிகவும் முக்கியமானது.
சான்றாதாரங்கள்
1. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. ஆக்ஸிஜனேற்ற அழுத்தத்தால் தூண்டப்பட்ட புற்றுநோயில் ஃப்ரீ ரேடிக்கல்கள், உலோகங்கள் மற்றும் ஆக்ஸிஜனேற்றிகள். கெம் பயோல் தொடர்பு. 2006;160:1-40.
2. ஹாலிவெல் பி, குட்டரிட்ஜ் ஜேஎம்சி. உயிரியல் மற்றும் மருத்துவத்தில் இலவச தீவிரவாதிகள். 3வது பதிப்பு. நியூயார்க்: ஆக்ஸ்போர்டு யுனிவர்சிட்டி பிரஸ்;1999.
3. மார்னெட் எல்ஜே. லிப்பிட் பெராக்ஸிடேஷன் மற்றும் மலோண்டியல்டிஹைடால் டிஎன்ஏ சேதம். முட்டாட் ரெஸ். 1999;424:83-95.
4. Siems WG, Grune T, Esterbauer H. 4-Hydroxynonenal உருவாக்கம் போது இஸ்கிமியா மற்றும் எலி சிறுகுடலின் மறுபிரவேசம். வாழ்க்கை அறிவியல். 1995;57:785-789.
5. ஸ்டாட்மேன் ஈஆர். வயதானதில் ஆக்ஸிஜனேற்ற இனங்களின் பங்கு. கர்ர் மெட் செம். 2004;11:1105-1112.
6. வாங் எம்ஒய், திங்ரா கே, ஹிட்டெல்மேன் டபிள்யூஎன், லீஹர் ஜேஜி, டிஆண்ட்ரேட் எம், லி டிஹெச். மனித மார்பக திசுக்களில் லிப்பிட் பெராக்ஸிடேஷன்-தூண்டப்பட்ட மலோண்டியல்டிஹைட் டிஎன்ஏ சேர்க்கிறது. புற்றுநோய் எபிடெமியோல் பயோமார்க்ஸ் முந்தைய 1996;5:705-710.
7. ஜென்னர் பி. பார்கின்சன் நோயில் ஆக்ஸிஜனேற்ற அழுத்தம். ஆன் நியூரோல். 2003;53: S26-S36.
8. லைராஸ் எல், கெய்ர்ன்ஸ் என்ஜே, ஜென்னர் ஏ, ஜென்னர் பி, ஹாலிவெல் பி ஜே நியூரோகெம். 1997;68:2061-2069.
9. சைர் எல்எம், ஸ்மித் எம்.ஏ., பெர்ரி ஜி. வேதியியல் மற்றும் நரம்பியக்கடத்தல் நோயில் ஆக்ஸிஜனேற்ற அழுத்தத்தின் உயிர்வேதியியல். கர்ர் மெட் செம். 2001;8:721-738.
10. தோஷ்னிவால் பிகே, ஸார்லிங் இஜே. மல்டிபிள் ஸ்களீரோசிஸில் லிப்பிட் பெராக்ஸைடேஷன் அதிகரித்ததற்கான சான்று. நியூரோகெம் ரெஸ். 1992;17:205-207.
11. Dhalla NS, Temsah RM, Netticadan T. இருதய நோய்களில் ஆக்ஸிஜனேற்ற அழுத்தத்தின் பங்கு. ஜே ஹைபர்டென்ஸ். 2000;18:655-673.
12. Kasparova S, Brezova V, Valko M, Horecky J, Mlynarik V, மற்றும் பலர். நாள்பட்ட மூளை ஹைப்போபெர்ஃபியூஷனின் எலி மாதிரியில் ஆக்ஸிஜனேற்ற அழுத்தத்தின் ஆய்வு. நியூரோகெம் இன்ட். 2005;46:601-611.
13. கெர் எஸ், ப்ரோஸ்னன் எம்ஜே, மெக்கின்டைர் எம், ரீட் ஜேஎல், டொமினிசாக் ஏஎஃப், ஹாமில்டன் சிஏ. மரபணு உயர் இரத்த அழுத்தத்தின் மாதிரியில் சூப்பர் ஆக்சைடு அயனி உற்பத்தி அதிகரிக்கப்படுகிறது: எண்டோடெலியத்தின் பங்கு. உயர் இரத்த அழுத்தம். 1999;33:1353-1358.
14. குக்ரேஜா ஆர்.சி., ஹெஸ் எம்.எல். ஆக்ஸிஜன் ஃப்ரீ-ரேடிக்கல் அமைப்பு: சமன்பாடுகளிலிருந்து சவ்வு-புரத தொடர்புகள் மூலம் இருதய காயம் மற்றும் பாதுகாப்பு வரை. கார்டியோவாஸ்க் ரெஸ். 1992;26:641-655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, மற்றும் பலர். சிகரெட் புகைத்தல் மனித நுரையீரலின் மையப் பகுதியில் 8-ஹைட்ராக்ஸிடாக்ஸிகுவானோசின் என்ற ஆக்ஸிஜனேற்ற டிஎன்ஏ பாதிப்பை அதிகரிக்கச் செய்கிறது. கார்சினோஜெனிசிஸ். 1997;18:1763-1766.
16. ஆண்ட்ரேடிஸ் ஏஏ, ஹேசன் எஸ்எல், கொஹேர் எஸ்ஏ, எர்சுரம் எஸ்சி. ஆஸ்துமாவில் ஆக்ஸிஜனேற்ற மற்றும் நைட்ரோசிட்டிவ் நிகழ்வுகள். இலவச ரேடிக் பயோல் மெட். 2003;35:213-225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, மற்றும் பலர். சிஸ்டமிக் சூப்பர் ஆக்சைடு டிஸ்முடேஸ் குறைபாடு ஆஸ்துமாவில் காற்று ஓட்டம் தடைபடும் தொடர்பு. ஆம் ஜே ரெஸ்பிர் கிரிட் கேர் மெட். 2005;172:306-313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, மற்றும் பலர். ஆஸ்துமா காற்றுப்பாதை மறுவடிவமைப்பு மற்றும் வினைத்திறன் நோய்க்குறியியல் இயற்பியலில் சூப்பர் ஆக்சைடு டிஸ்முடேஸ் செயலிழப்பு. ஆம் ஜே பதோல். 2005;166:663-674.
19. Dut R, Dizdar EA, Birben E, Sackesen C, Soyer OU, Besler T, Kalayci O. ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் ஆஸ்துமா உள்ள குழந்தைகளின் காற்றுப்பாதைகளில் அதன் தீர்மானங்கள். ஒவ்வாமை. 2008;63:1605-1609.
20. எர்கன் எச், பிர்பென் ஈ, டிஜ்டார் ஈஏ, கெஸ்கின் ஓ, கராஸ்லான் சி மற்றும் பலர். குழந்தை பருவ ஆஸ்துமாவில் ஆக்ஸிஜனேற்ற காயத்தின் ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் மரபியல் மற்றும் தொற்றுநோயியல் தீர்மானிப்பவர்கள். ஜே அலர்ஜி கிளினின் இம்யூனோல். 2006;118:1097-1104.
21. Fitzpatrick AM, Teague WG, Holguin F, Yeh M, Brown LA. கடுமையான ஆஸ்துமா ஆராய்ச்சி திட்டம். கடுமையான ஆஸ்துமா உள்ள குழந்தைகளில் ஏர்வே குளுதாதயோன் ஹோமியோஸ்டாஸிஸ் மாற்றப்படுகிறது: ஆக்ஸிஜனேற்ற அழுத்தத்திற்கான ஆதாரம். ஜே அலர்ஜி கிளினின் இம்யூனோல். 2009;123:146-152.
22. மில்லர் டிஎம், ப்யூட்னர் ஜிஆர், ஆஸ்ட் எஸ்டி. "ஆக்ஸிடேஷன்" எதிர்வினைகளின் வினையூக்கிகளாக மாற்ற உலோகங்கள். இலவச ரேடிக் பயோல் மெட். 1990;8:95-108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, D'me D, Pommier J. தைராய்டு பிளாஸ்மா மென்படலத்தில் NADPH ஆக்சிடேஸ் மூலம் வினையூக்கி ஹைட்ரஜன் பெராக்சைடு உருவாக்கம். ஜே பயோல் கெம். 1991;266:3739-3743.
24. கிரேஞ்சர் டிஎன். இஸ்கிமியர்பெர்ஃபியூஷன் காயத்தில் சாந்தைன் ஆக்சிடேஸ் மற்றும் கிரானுலோசைட்டுகளின் பங்கு. ஆம் ஜே பிசியோல். 1988;255:H1269-H1275.
25. ஃபென்டன் HJH. இரும்பு முன்னிலையில் டார்டாரிக் அமிலத்தின் ஆக்சிஜனேற்றம். J Chem Soc. 1984;65:899-910.
26. ஹேபர் எஃப், வெயிஸ் ஜே.ஜே. இரும்பு உப்புகளால் ஹைட்ரஜன் பெராக்சைட்டின் வினையூக்க சிதைவு. Proc R Soc Lond Ser A. 1934;147:332-351.
27. லியோசெவ் எஸ்ஐ, ஃப்ரிடோவிச் I. ஹேபர் வெயிஸ் 70 ஆண்டுகளுக்குப் பிறகு சைக்கிள் ஓட்டினார்: ஒரு மாற்றுக் கருத்து. ரெடாக்ஸ் ரெப். 2002;7:55-57.
28. கிளெபனாஃப் எஸ்.ஜே. Myeloperoxidase: நண்பர் மற்றும் எதிரி. ஜே லுகோக் பயோல். 2005;77:598-625.
29. ஒயிட்மேன் எம், ஜென்னர் ஏ, ஹாலிவெல் பி. தனிமைப்படுத்தப்பட்ட கன்று தைமஸ் டிஎன்ஏவில் ஹைபோகுளோரஸ் அமிலத்தால் தூண்டப்பட்ட அடிப்படை மாற்றங்கள். கெம் ரெஸ் டாக்ஸிகோல். 1997;10:1240-1246.
30. Kulcharyk PA, Heinecke JW. மனித பாகோசைட்டுகளின் மைலோபெராக்ஸிடேஸ் அமைப்பால் உற்பத்தி செய்யப்படும் ஹைப்போகுளோரஸ் அமிலம் டிஎன்ஏ மற்றும் புரதங்களுக்கு இடையே கோவலன்ட் குறுக்கு இணைப்புகளைத் தூண்டுகிறது. உயிர்வேதியியல். 2001;40:3648–3656.
31. பிரென்னன் எம்எல், வூ டபிள்யூ, ஃபூ எக்ஸ், ஷென் இசட், சாங் டபிள்யூ, மற்றும் பலர். இரண்டு சர்ச்சைகளின் கதை: ஈசினோபில் பெராக்ஸிடேஸ் மற்றும் மைலோபெராக்ஸிடேஸ் குறைபாடுள்ள எலிகளைப் பயன்படுத்தி விவோவில் நைட்ரோடைரோசின் உருவாவதில் பெராக்ஸிடேஸின் பங்கு மற்றும் பெராக்ஸிடேஸ்-உருவாக்கும் எதிர்வினை நைட்ரஜன் இனங்களின் தன்மை ஆகிய இரண்டையும் வரையறுத்தல். ஜே பயோல் கெம். 2002;277:17415-17427.
32. Denzler KL, Borchers MT, Crosby JR, Cieslewicz G, Hines EM, மற்றும் பலர். நுரையீரல் அழற்சியின் மவுஸ் ஓவல்புமின்-சவால் மாதிரியில் விரிவான ஈசினோபில் சிதைவு மற்றும் காற்றுப்பாதை புரதங்களின் பெராக்ஸிடேஸ்-மத்தியஸ்த ஆக்சிஜனேற்றம் ஏற்படாது. ஜே இம்யூனோல். 2001;167:1672-1682.
33. வான் டேலன் CJ, Winterbourn CC, செந்தில்மோகன் R, கெட்டில் AJ. நைட்ரைட் ஒரு அடி மூலக்கூறு மற்றும் மைலோபெராக்ஸிடேஸின் தடுப்பானாக. அழற்சியின் இடங்களில் நைட்ரேஷன் மற்றும் ஹைபோகுளோரஸ் அமில உற்பத்திக்கான தாக்கங்கள். ஜே பயோல் கெம். 2000;275:11638-11644.
34. வூட் எல்ஜி, ஃபிட்ஸ்ஜெரால்ட் டிஏ, கிப்சன் பிஜி, கூப்பர் டிஎம், கார்க் எம்எல். பிளாஸ்மா ஐசோபிரோஸ்டேன்களால் தீர்மானிக்கப்படும் லிப்பிட் பெராக்சிடேஷன் லேசான ஆஸ்துமாவின் நோயின் தீவிரத்துடன் தொடர்புடையது. லிப்பிடுகள். 2000;35:967-974.
35. Montuschi P, Corradi M, Ciabattoni G, Nightingale J, Kharitonov SA, Barnes PJ. ஆஸ்துமா நோயாளிகளின் வெளியேற்றப்பட்ட மின்தேக்கியில், ஆக்ஸிஜனேற்ற அழுத்தத்தின் குறிப்பானான 8-ஐசோப்ரோஸ்டேன் அதிகரித்தது. ஆம் ஜே ரெஸ்பிர் கிரிட் கேர் மெட். 1999;160:216-220.
36. சர்ச் DF, பிரையர் WA. சிகரெட் புகையின் ஃப்ரீ-ரேடிக்கல் வேதியியல் மற்றும் அதன் நச்சுயியல் தாக்கங்கள். சுற்றுச்சூழல் சுகாதார பார்வை. 1985;64:111-126.
37. Hiltermann JT, Lapperre TS, van Bree L, Steerenberg PA, Brahim JJ, et al. ஆஸ்துமா நோயாளிகளின் ஸ்பூட்டம் மற்றும் மூச்சுக்குழாய் கழுவும் திரவத்தில் ஓசோன் தூண்டப்பட்ட அழற்சி மதிப்பிடப்படுகிறது: காற்று மாசுபாடு மற்றும் ஆஸ்துமா பற்றிய தொற்றுநோயியல் ஆய்வுகளில் ஒரு புதிய ஆக்கிரமிப்பு அல்லாத கருவி. இலவச ரேடிக் பயோல் மெட். 1999;27:1448-1454.
38. நைட்டிங்கேல் ஜேஏ, ரோஜர்ஸ் டிஎஃப், பார்ன்ஸ் பிஜே. சாதாரண மற்றும் ஆஸ்துமா நோயாளிகளில் வெளியேற்றப்படும் நைட்ரிக் ஆக்சைடு, நுரையீரல் செயல்பாடு மற்றும் தூண்டப்பட்ட சளி ஆகியவற்றில் உள்ளிழுக்கப்படும் ஓசோனின் விளைவு. தோராக்ஸ். 1999;54:1061-1069.
39. சோ AK, Sioutas C, Miguel AH, Kumagai Y, Schmitz DA, மற்றும் பலர். லாஸ் ஏஞ்சல்ஸ் பேசின் வெவ்வேறு தளங்களில் காற்றில் பரவும் துகள்களின் ரெடாக்ஸ் செயல்பாடு. சுற்றுச்சூழல் Res. 2005;99:40-47.
40. Comhair SA, Thomassen MJ, Erzurum SC. 2% O(100) அல்லது சிகரெட் புகையால் வெளிப்படும் ஆரோக்கியமான நபர்களின் காற்றுப்பாதைகளில் எக்ஸ்ட்ராசெல்லுலர் குளுதாதயோன் பெராக்சிடேஸ் மற்றும் நைட்ரிக் ஆக்சைடு சின்தேஸ் 2 ஆகியவற்றின் மாறுபட்ட தூண்டல். ஆம் ஜே ரெஸ்பிர் செல் மோல் பயோல். 2000;23:350–354.
41. மத்தே எம்.ஏ., கீசர் டி, மாடலோன் எஸ், இஸ்கிரோபொலோஸ் எச். ஆக்சிடன்ட்-மத்தியஸ்த நுரையீரல் காயம் கடுமையான சுவாசக் கோளாறு நோய்க்குறி. கிரிட் கேர் மெட். 1999;27:2028-2030.
42. Biaglow JE, Mitchell JB, Hold K. X-ray பதிலில் பெராக்சைடு மற்றும் சூப்பர் ஆக்சைட்டின் முக்கியத்துவம். இன்ட் ஜே ரேடியட் ஒன்கோல் பயோல் பிசிஸ். 1992;22:665-669.
43. Chiu SM, Xue LY, Friedman LR, Oleinick NL. அயனியாக்கும் கதிர்வீச்சுக்கு அணுக்கரு அணி இணைப்புத் தளங்களின் செப்பு அயனி-மத்தியஸ்த உணர்திறன். உயிர்வேதியியல். 1993;32:6214-6219.
44. நாராயணன் பிகே, குட்வின் இஎச், லெஹ்னெர்ட் பிஇ. ஆல்பா துகள்கள் மனித உயிரணுக்களில் சூப்பர் ஆக்சைடு அனான்கள் மற்றும் ஹைட்ரஜன் பெராக்சைடு ஆகியவற்றின் உயிரியல் உற்பத்தியைத் தொடங்குகின்றன. புற்றுநோய் ரெஸ். 1997;57:3963-3971.
45. டட்டில் SW, வார்னஸ் ME, மிட்செல் JB, Biaglow JE. ஆக்ஸிஜனேற்ற பென்டோஸ் சுழற்சி செயல்பாட்டில் குறைபாடுள்ள CHO செல் கோடுகளில் இரசாயன ஆக்ஸிஜனேற்றங்கள் மற்றும் கதிர்வீச்சுக்கான உணர்திறன். இன்ட் ஜே ரேடியட் ஓன்கோல் பயோல் பிசிஸ். 1992;22: 671-675.
46. Guo G, Yan-Sanders Y, Lyn-Cook BD, Wang T, Tamae D, மற்றும் பலர். மாங்கனீசு
கதிர்வீச்சினால் தூண்டப்பட்ட சூப்பர் ஆக்சைடு டிஸ்முடேஸ்-மத்தியஸ்த மரபணு வெளிப்பாடு
தகவமைப்பு பதில்கள். மோல் செல் பயோல். 2003;23:2362-2378.
47. Azzam EI, de Toledo SM, Spitz DR, Little JB. ஆக்ஸிஜனேற்ற வளர்சிதை மாற்றம்
சிக்னல் கடத்தல் மற்றும் பார்வையாளர்களில் மைக்ரோநியூக்ளியஸ் உருவாக்கத்தை மாற்றியமைக்கிறது
ஒரு-துகள் கதிர்வீச்சு சாதாரண மனித ஃபைப்ரோபிளாஸ்ட்களில் இருந்து செல்கள். புற்றுநோய் ரெஸ்.
2002;62:5436–5442.
48. லீச் ஜேகே, வான் டுய்ல் ஜி, லின் பிஎஸ், ஷ்மிட்-உல்ரிச் ஆர், மிக்கெல்சன் ஆர்பி.
அயனியாக்கும் கதிர்வீச்சினால் தூண்டப்பட்ட, மைட்டோகாண்ட்ரியா சார்ந்த வினைத்திறன்
ஆக்ஸிஜன் / நைட்ரஜன். புற்றுநோய் ரெஸ். 2001;61:3894-3901.
49. Dent P, Yacoub A, Fisher PB, Hagan MP, Grant S. MAPK பாதைகள் உள்ள
கதிர்வீச்சு பதில்கள். புற்றுநோயியல். 2003;22:5885–5896.
50. வெய் எஸ்ஜே, போடெரோ ஏ, ஹிரோடா கே, பிராட்பரி சிஎம், மார்கோவினா எஸ், மற்றும் பலர். தியோரெடாக்சின்
அணுக்கரு இடமாற்றம் மற்றும் ரெடாக்ஸ் காரணி-1 உடனான தொடர்பு அயனியாக்கும் கதிர்வீச்சுக்கு பதிலளிக்கும் வகையில் AP-1 டிரான்ஸ்கிரிப்ஷன் காரணியை செயல்படுத்துகிறது. புற்றுநோய் ரெஸ். 2000;60:6688–6695.
51. கேடட் ஜே, டௌகி டி, காஸ்பருட்டோ டி, ராவணத் ஜேஎல். டிஎன்ஏவுக்கு ஆக்ஸிஜனேற்ற சேதம்: உருவாக்கம், அளவீடு மற்றும் உயிர்வேதியியல் அம்சங்கள். முட்டாட் ரெஸ். 2003;531:5-23.
52. Yokoya A, Cunniffe SM, O'Neill P. காமரேடியேஷன் மூலம் பிளாஸ்மிட் டிஎன்ஏ படங்களில் ஸ்ட்ராண்ட் பிரேக்ஸ் மற்றும் பேஸ் புண்கள் தூண்டுதலின் மீது நீரேற்றத்தின் விளைவு. ஜே ஆம் கெம் சோக். 2002;124:8859–8866.
53. ஜான்சென் ஒய்எம், வான் ஹவுடன் பி, போர்ம் பிஜே, மோஸ்மேன் பிடி. ஆக்ஸிஜனேற்ற சேதத்திற்கு செல் மற்றும் திசு பதில்கள். ஆய்வக முதலீடு. 1993;69:261-274.
54. இவனகா எம், மோரி கே, ஐடா டி, உராடா ஒய், மாட்சுவோ டி, மற்றும் பலர். T98G மனித கிளியோபிளாஸ்டோமா செல்களில் அயனியாக்கும் கதிர்வீச்சு மூலம் காமா குளுட்டமைல்சிஸ்டைன் சின்தேடேஸின் அணுக்கரு காரணி கப்பா பி சார்ந்த தூண்டல். இலவச ரேடிக் பயோல் மெட். 1998;24:1256-1268.
55. Stohs SJ, Bagchi D. உலோக அயனிகளின் நச்சுத்தன்மையில் ஆக்ஸிஜனேற்ற வழிமுறைகள். இலவச ரேடிக் பயோல் மெட். 1995;18:321-336.
56. லியோனார்ட் எஸ்எஸ், ஹாரிஸ் ஜிகே, ஷி எக்ஸ். உலோகத்தால் தூண்டப்பட்ட ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் சமிக்ஞை கடத்தல். இலவச ரேடிக் பயோல் மெட். 2004;37:1921-1942.
57. ஷி எச், ஷி எக்ஸ், லியு கேஜே. ஆர்சனிக் நச்சுத்தன்மை மற்றும் புற்றுநோயின் ஆக்ஸிஜனேற்ற வழிமுறை. மோல் செல் உயிர்வேதியியல். 2004;255:67-78.
58. பை ஜே, ஹோரிகுச்சி எஸ், சன் ஒய், நிகைடோ எம், ஷிமோஜோ என், ஹயாஷி டி. இலவச ரேடிக் பயோல் மெட்.2003;35:102-113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, Okada S. டிமெதிலார்சினிக் அமிலத்தால் தூண்டப்பட்ட டிஎன்ஏஸ்ட்ராண்ட் முறிவுகள், கனிம ஆர்சனிக்ஸின் வளர்சிதை மாற்றமானது, சூப்பர் ஆக்சைடு அயனி ரேடிக்கல்களால் வலுவாக மேம்படுத்தப்படுகிறது. பயோல் பார்ம் புல். 1995;18:45-58.
60. வால்கேஸ் எம்பி, லியு ஜே, வார்டு ஜேஎம், திவான் எல்ஏ. ஆர்சனிக் கார்சினோஜெனிசிஸ் அடிப்படையிலான வழிமுறைகள்: கர்ப்ப காலத்தில் கனிம ஆர்சனிக் வெளிப்படும் எலிகளின் அதிக உணர்திறன். நச்சுயியல். 2004;198:31-38.
61. ஷில்லர் சிஎம், ஃபோலர் பிஏ, வூட்ஸ் ஜேஎஸ். பைருவேட் டீஹைட்ரோஜினேஸ் செயல்பாட்டில் ஆர்சனிக்கின் விளைவுகள். சுற்றுச்சூழல் சுகாதார பார்வை. 1977;19:205-207.
62. Monterio HP, Bechara EJH, அப்தல்லா DSP. நரம்பியல் போர்பிரியாஸ் மற்றும் ஈய விஷத்தில் ஃப்ரீ ரேடிக்கல்களின் ஈடுபாடு. மோல் செல் உயிர்வேதியியல். 1991;103:73-83.
63. திரிபாதி ஆர்எம், ரகுநாத் ஆர், மஹாபத்ரா எஸ். இரத்த ஈயம் மற்றும் குழந்தைகளின் சிடி, கியூ, ஸிஎன், ஃபெ மற்றும் ஹீமோகுளோபின் அளவுகளில் அதன் விளைவு. அறிவியல் மொத்த சூழல். 2001;277:161-168.
64. நேரு பி, துவா ஆர். ஈய நியூரோடாக்சிசிட்டியில் உணவு செலினியத்தின் விளைவு. ஜே சுற்றுச்சூழல் பாத்தோல் டாக்ஸிகால் ஓன்கோல். 1997;16:47-50.
65. ரீட் டிஎம், ஃபீக் டிஐ, லோப் எல்ஏ. உலோகத்தால் தூண்டப்பட்ட ஆக்ஸிஜன் ரேடிக்கல்களால் பிறழ்வு. சுற்றுச்சூழல் சுகாதார பார்வை. 1994;102(suppl 3):57–61.
66. கின்னுலா வி.எல், க்ராபோ ஜே.டி. நுரையீரல் மற்றும் மனித நுரையீரல் நோய்களில் சூப்பர் ஆக்சைடு டிஸ்முடேஸ்கள். ஆம் ஜே ரெஸ்பிர் கிரிட் கேர் மெட். 2003;167:1600-1619.
67. கின்னுலா வி.எல். மனித நுரையீரலில் ஏற்படும் அழற்சி நிலைகளின் போது ஆக்ஸிஜன் வளர்சிதை மாற்றங்களின் உற்பத்தி மற்றும் சிதைவு. கர்ர் மருந்து அழற்சி ஒவ்வாமையை குறிவைக்கிறது. 2005;4:465-470.
68. Zelko IN, மரியானி TJ, Folz RJ. Superoxide dismutase multigene குடும்பம்: CuZn-SOD (SOD1), Mn-SOD (SOD2), மற்றும் EC-SOD (SOD3) மரபணு கட்டமைப்புகள், பரிணாமம் மற்றும் வெளிப்பாடு ஆகியவற்றின் ஒப்பீடு. இலவச ரேடிக் பயோல் மெட். 2002;33:337-349.
69. கிர்க்மேன் எச்என், ரோல்ஃபோ எம், ஃபெராரிஸ் ஏஎம், கெடானி ஜிஎஃப். என்ஏடிபிஎச் மூலம் கேடலேஸின் பாதுகாப்பு வழிமுறைகள். இயக்கவியல் மற்றும் ஸ்டோச்சியோமெட்ரி. ஜே பயோல் கெம். 1999;274:13908-13914.
70. ஃப்ளோஹோ எல். குளுதாதயோன் பெராக்ஸிடேஸ். அடிப்படை வாழ்க்கை அறிவியல். 1988;49:663-668.
71. ஆர்தர் ஜே.ஆர். குளுதாதயோன் பெராக்ஸிடேஸ்கள். செல் மோல் லைஃப் அறிவியல். 2000;57:1825-1835.
72. சூ எஃப்எஃப், டோரோஷோ ஜேஎச், எஸ்வர்தி ஆர்எஸ். புதிய செல்லுலார் செலினியம் சார்ந்த குளுதாதயோன் பெராக்ஸிடேஸின் வெளிப்பாடு, குணாதிசயம் மற்றும் திசு விநியோகம், GSHPx-GI. ஜே பயோல் கெம். 1993;268:2571-2576.
73. Comhair SA, Bathena PR, Farver C, Thunnissen FB, Erzurum SC. ஆஸ்துமா நுரையீரலில் எக்ஸ்ட்ராசெல்லுலர் குளுதாதயோன் பெராக்ஸிடேஸ் தூண்டல்: மனித சுவாசப்பாதை எபிடெலியல் செல்களில் வெளிப்பாட்டின் ரெடாக்ஸ் ஒழுங்குமுறைக்கான சான்று. FASEB ஜே. 2001;15:70-78.
74. க்ரோமர் எஸ், யூரிக் எஸ், பெக்கர் கே. தியோரெடாக்சின் அமைப்பு அறிவியலில் இருந்து கிளினிக் வரை. மெட் ரெஸ் ரெவ். 2004;24:40-89.
75. Kinnula VL, Lehtonen S, Kaarteenaho-Wiik R, Lakari E, P''kk' P, et al. மனித நுரையீரல் மற்றும் நுரையீரல் சார்கோயிடோசிஸில் பெராக்ஸிரெடாக்ஸின்களின் செல் குறிப்பிட்ட வெளிப்பாடு. தோராக்ஸ். 2002;57:157-164.
76. Dubuisson M, Vander Stricht D, Clippe A, Etienne F, Nauser T, மற்றும் பலர். மனித பெராக்சிரெடாக்சின் 5 என்பது பெராக்ஸைனிட்ரைட் ரிடக்டேஸ் ஆகும். FEBS லெட். 2004;571:161-165.
77. ஹோல்ம்கிரென் ஏ. தியோரெடாக்சின் மற்றும் குளுடரெடாக்சின் அமைப்புகளின் ஆக்ஸிஜனேற்ற செயல்பாடு. ஆன்டிஆக்சிட் ரெடாக்ஸ் சிக்னல். 2000;2:811-820.
78. டிக்கின்சன் டிஏ, ஃபார்மன் எச்.ஜே. பாதுகாப்பு மற்றும் சமிக்ஞைகளில் குளுதாதயோன்: ஒரு சிறிய தியோலில் இருந்து பாடங்கள். Ann NY Acad Sci. 2002;973:488-504.
79. Sies H. குளுதாதயோன் மற்றும் செல்லுலார் செயல்பாடுகளில் அதன் பங்கு. இலவச ரேடிக் பயோல் மெட். 1999;27:916-921.
80. லாட்னர் ஜேஇ, பார்சன்ஸ் ஜேஎஃப், ரைஃப் சிஎல், கில்லிலேண்ட் ஜிஎல், ஆம்ஸ்ட்ராங் ஆர்என். குளுதாதயோன் பரிமாற்றங்களுக்கான இணையான பரிணாம பாதைகள்: மைட்டோகாண்ட்ரியல் கிளாஸ் கப்பா என்சைம் rGSTK1-1 இன் அமைப்பு மற்றும் வழிமுறை. உயிர்வேதியியல். 2004;43:52-61.
81. ராபின்சன் ஏ, ஹட்லி ஜிஏ, பூத் எச்எஸ், போர்டு பிஜி. மனித கப்பா கிளாஸ் குளுதாதயோன் டிரான்ஸ்ஃபரேஸின் மாடலிங் மற்றும் பயோ-இன்ஃபர்மேடிக்ஸ் ஆய்வுகள், புரோகாரியோடிக் 2-ஹைட்ராக்ஸிக்ரோமீன்-2-கார்பாக்சிலேட் ஐசோமரேஸ்களுக்கு ஹோமோலஜியுடன் ஒரு புதுமையான மூன்றாவது டிரான்ஸ்ஃபரேஸ் குடும்பத்தைக் கணிக்கின்றன. பயோகெம் ஜே. 2004;379:541–552.
82. Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson A, Persson B. ஐகோசனாய்டு மற்றும் குளுதாதயோன் வளர்சிதை மாற்றத்தில் மிகவும் மாறுபட்ட செயல்பாடுகளுடன் கூடிய சவ்வு தொடர்புடைய புரதங்களின் பரவலான சூப்பர் குடும்பமான MAPEGda இன் பொதுவான கட்டமைப்பு அம்சங்கள். புரதம் அறிவியல். 1999;8:689-692.
83. ஹேய்ஸ் ஜேடி, புல்ஃபோர்ட் டிஜே. குளுதாதயோன் எஸ்-டிரான்ஸ்ஃபெரேஸ் சூப்பர்ஜீன் குடும்பம்: ஜிஎஸ்டியின் கட்டுப்பாடு மற்றும் புற்றுநோய் வேதியியல் பாதுகாப்பு மற்றும் மருந்து எதிர்ப்பு ஆகியவற்றில் ஐசோஎன்சைம்களின் பங்களிப்பு. Crit Rev Biochem Mol Biol. 1995;30:445-600.
84. ஆம்ஸ்ட்ராங் ஆர்என். குளுதாதயோன் பரிமாற்றங்களின் அமைப்பு, வினையூக்கி பொறிமுறை மற்றும் பரிணாமம். கெம் ரெஸ் டாக்ஸிகோல். 1997;10:2-18.
85. ஹேய்ஸ் ஜேடி, மெக்லெலன் எல்ஐ. குளுதாதயோன் மற்றும் குளுதாதயோன் சார்ந்த நொதிகள் ஆக்ஸிஜனேற்ற அழுத்தத்திற்கு எதிராக ஒருங்கிணைந்த ஒழுங்குபடுத்தப்பட்ட பாதுகாப்பைக் குறிக்கின்றன. இலவச ரேடிக் ரெஸ். 1999;31:273-300.
86. ஷீஹான் டி, மீடே ஜி, ஃபோலே விஎம், டவுட் சிஏ. குளுதாதயோன் பரிமாற்றங்களின் அமைப்பு, செயல்பாடு மற்றும் பரிணாமம்: ஒரு பண்டைய நொதி சூப்பர் குடும்பத்தின் பாலூட்டி அல்லாத உறுப்பினர்களை வகைப்படுத்துவதற்கான தாக்கங்கள். பயோகெம் ஜே. 2001;360:1-16.
87. சோ எஸ்ஜி, லீ ஒய்எச், பார்க் எச்எஸ், ரியோ கே, காங் கேடபிள்யூ, மற்றும் பலர். Glutathione S-transferase Mu ஆனது அப்போப்டொசிஸ் சிக்னல்-ஒழுங்குபடுத்தும் கைனேஸ் 1. J Biol Chem ஐ அடக்குவதன் மூலம் அழுத்தம் செயல்படுத்தப்பட்ட சமிக்ஞைகளை மாற்றியமைக்கிறது. 2001;276:12749-12755.
88. டோரியன் எஸ், லம்பேர்ட் எச், லேண்ட்ரி ஜே ஜே பயோல் கெம். 38;1:2002-277.
89. அட்லர் V, யின் Z, Fuchs SY, Benezra M, Rosario L, மற்றும் பலர். ஜிஎஸ்டிபி மூலம் ஜேஎன்கே சிக்னலை ஒழுங்குபடுத்துதல். எம்போ ஜே. 1999;18:1321-1334.
90. மனேவிச் ஒய், ஃபைன்ஸ்டீன் எஸ்ஐ, ஃபிஷர் ஏபி. ஆக்ஸிஜனேற்ற நொதி 1-CYS பெராக்ஸிரெடாக்சின் செயல்படுத்துவதற்கு pGST உடன் ஹீட்டோரோடைமரைசேஷன் மூலம் மத்தியஸ்தம் செய்யப்பட்ட குளுதாதயோனைலேஷன் தேவைப்படுகிறது. Proc Natl Acad Sci US A. 2004;101:3780–3785.
91. பங்கர் VW. ஃப்ரீ ரேடிக்கல்கள், ஆக்ஸிஜனேற்றிகள் மற்றும் வயதானவர்கள். மெட் லேப் அறிவியல். 1992;49:299-312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD, மற்றும் பலர். முறையான ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் வயது மற்றும் நோயுடனான அதன் உறவு. ஜே ஆம் ஜெரியாட்டர் சோக். 1996;44:823-827.
93. ஒயிட் இ, ஷானன் ஜேஎஸ், பேட்டர்சன் ஆர்.ஈ. வைட்டமின் மற்றும் இடையே உள்ள உறவு
கால்சியம் கூடுதல் பயன்பாடு மற்றும் பெருங்குடல் புற்றுநோய். புற்றுநோய் எபிடெமியோல் பயோமார்க்ஸ் முந்தைய 1997;6:769-774.
94. Masella R, Di Benedetto R, Vari R, Filesi C, Giovannini C. உயிரியல் அமைப்புகளில் இயற்கை ஆக்ஸிஜனேற்ற கலவைகளின் நாவல் வழிமுறைகள்: குளுதாதயோன் மற்றும் குளுதாதயோன் தொடர்பான நொதிகளின் ஈடுபாடு. ஜே நியூட்ர் உயிர்வேதியியல். 2005;16:577-586.
95. குரெல்லோ எஸ், செகோனி சி, பிகோலி சி, ஃபெராரி ஆர், ஆல்பர்டினி ஏ, குர்னியேரி சி அனுபவம். 1985;41:42-43.
96. எல்-அகாமி ஏ, லோவ் ஜிஎம், மெக்கார்வி டிஜே, மோர்டென்சன் ஏ, பிலிப் டிஎம், டிரஸ்கோட் டிஜி. கரோட்டினாய்டு தீவிர வேதியியல் மற்றும் ஆக்ஸிஜனேற்ற/சார்பு-ஆக்ஸிடன்ட் பண்புகள். Arch Biochem Biophys. 2004;430:37-48.
97. ரைஸ்-எவன்ஸ் CA, சாம்ப்சன் ஜே, பிராம்லி PM, ஹாலோவே DE. விவோவில் கரோட்டினாய்டுகள் ஆக்ஸிஜனேற்றிகளாக இருக்கும் என்று நாம் ஏன் எதிர்பார்க்கிறோம்? இலவச ரேடிக் ரெஸ். 1997;26:381-398.
98. நைல்ஸ் ஆர்.எம். ரெட்டினாய்டு வேதியியல் தடுப்பு மற்றும் புற்றுநோய் சிகிச்சையில் சிக்னலிங் பாதைகள். முட்டாட் ரெஸ். 2004;555:81-96.
99. டோனாடோ எல்ஜே, நோய் என். ரெட்டினோயிக் அமிலத்தால் மார்பகப் புற்றுநோய் வளர்ச்சியை அடக்குதல்: ப்ரோபொப்டோடிக் மரபணுக்கள் ரெட்டினோயிக் அமிலம் ஏற்பி மற்றும் செல்லுலார் ரெட்டினோயிக் அமிலம்-பிணைப்பு புரதம் II சமிக்ஞைக்கான இலக்குகளாகும். புற்றுநோய் ரெஸ். 2005;65:8193-8199.
100. Niizuma H, Nakamura Y, Ozaki T, Nakanishi H, Ohira M, et al. Bcl-2 என்பது நியூரோபிளாஸ்டோமாவில் ரெட்டினோயிக் அமிலத்தால் தூண்டப்பட்ட அப்போப்டொடிக் செல் இறப்பிற்கான முக்கிய சீராக்கி ஆகும். புற்றுநோயியல். 2006;25:5046–5055.
101. டால்டன் TP, ஷெர்ட்சர் HG, Puga A. எதிர்வினை ஆக்ஸிஜன் மூலம் மரபணு வெளிப்பாட்டைக் கட்டுப்படுத்துதல். ஆன் ரெவ் பார்மகோல் டாக்ஸிகோல். 1999;39:67-101.
102. ஸ்காண்டலியோஸ் ஜே.ஜி. ஆக்ஸிஜனேற்ற அழுத்தத்திற்கான மரபணு பதில்கள். இல்: மேயர்ஸ் RA, எட். மூலக்கூறு உயிரணு உயிரியல் மற்றும் மூலக்கூறு மருத்துவத்தின் கலைக்களஞ்சியம். தொகுதி 5. 2வது பதிப்பு. Weinheim, ஜெர்மனி: Wiley-VCH; 2004: 489–512.
103. கோஷ் ஆர், மிட்செல் டிஎல். டிரான்ஸ்கிரிப்ஷன் காரணி பிணைப்பில் ஊக்குவிப்பு உறுப்புகளில் ஆக்ஸிஜனேற்ற டிஎன்ஏ சேதத்தின் விளைவு. நியூக்ளிக் அமிலங்கள் ரெஸ். 1999;27:3213-3218.
104. மரியெட்டா சி, குலாம் எச், ப்ரூக்ஸ் பிஜே. TATA பெட்டியில் உள்ள ஒரு ஒற்றை 8, 50-cyclo-20-deoxyadenosine புண் TATA பிணைப்பு புரதத்தை பிணைப்பதைத் தடுக்கிறது மற்றும் விவோவில் டிரான்ஸ்கிரிப்ஷனை வலுவாகக் குறைக்கிறது. டிஎன்ஏ பழுதுபார்ப்பு (ஆம்ஸ்ட்). 2002;1:967-975.
105. ஜாக்சன் AL, சென் ஆர், லோப் LA. மைக்ரோசாட்லைட் உறுதியற்ற தன்மையின் தூண்டல்
ஆக்ஸிஜனேற்ற டிஎன்ஏ சேதத்தால். Proc Natl Acad Sci US A. 1998;95:12468–12473.
106. கால்டெகாட் KW. பாலூட்டிகளின் டிஎன்ஏ ஒற்றை இழை முறிவு பழுதுபார்க்கும் போது புரதம்-புரத தொடர்பு. பயோகெம் Soc Trans. 2003;31:247-251.
107. குக் எம்.எஸ்., எவன்ஸ் எம்.டி., டிஸ்டாரோக்லு எம்., லுனெக் ஜே. ஆக்ஸிஜனேற்ற டிஎன்ஏ சேதம்: வழிமுறைகள், பிறழ்வு மற்றும் நோய். FASEB ஜே. 2003;17:1195-1214.
108. ஜோன்ஸ் PL, Wolffe AP. மரபணு வெளிப்பாட்டைத் தீர்மானிப்பதில் குரோமாடின் அமைப்பு மற்றும் டிஎன்ஏ மெத்திலேஷன் ஆகியவற்றுக்கு இடையேயான உறவுகள். Semin Cancer Biol. 1999;9:339-347.
109. ஜிரோட்டி AW. லிப்பிட் பெராக்ஸைடேஷன் வழிமுறைகள். ஜே ஃப்ரீ ரேடிக் பயோல் மெட். 1985;1:87-95.
110. சியு ஜிஎம், டிராப்பர் ஹெச்எச். விவோ மற்றும் இன் விட்ரோவில் மலோனால்டிஹைட்டின் வளர்சிதை மாற்றம். லிப்பிடுகள். 1982;17:349-355.
111. Esterbauer H, Koller E, Slee RG, Koster JF. ஃப்ளோரசன்ட் குரோமோலிப்பிட்களின் உருவாக்கத்தில் லிப்பிட்-பெராக்ஸிடேஷன் தயாரிப்பு 4-ஹைட்ராக்ஸினோனெனல் சாத்தியமான ஈடுபாடு. பயோகெம் ஜே. 1986;239:405-409.
112. Hagihara M, Nishigaki I, Maseki M, Yagi K. மனித சீரம் லிப்போபுரோட்டீன் பின்னங்களில் லிப்பிட் பெராக்சைடு அளவுகளில் வயது சார்ந்த மாற்றங்கள். ஜே ஜெரண்டோல். 1984;39:269-272.
113. கெல்லர் ஜேஎன், மார்க் ஆர்ஜே, புரூஸ் ஏஜே, பிளாங்க் ஈ, ரோத்ஸ்டீன் ஜேடி, மற்றும் பலர். 4- ஹைட்ராக்ஸினோனெனல், சவ்வு லிப்பிட் பெராக்சிடேஷனின் ஆல்டிஹைடிக் தயாரிப்பு, சினாப்டோசோம்களில் குளுட்டமேட் போக்குவரத்து மற்றும் மைட்டோகாண்ட்ரியல் செயல்பாட்டை பாதிக்கிறது. நரம்பியல். 1997;806:85-96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. லிப்பிட் பெராக்சிடேஷனின் இறுதிப் பொருளின் மூலம் அழுத்த சமிக்ஞை பாதைகளை செயல்படுத்துதல். 4-ஹைட்ராக்ஸி-2-நோனெனல் என்பது உயிரணுக்களுக்குள் பெராக்சைடு உற்பத்தியின் சாத்தியமான தூண்டியாகும். ஜே பயோல் கெம். 1999;274:2234-2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. ஆக்ஸிஜனேற்றப்பட்ட LDL மூலம் EGF ஏற்பியை செயல்படுத்துதல். FASEB ஜே. 1998;12:665-671.
116. Tsukagoshi H, Kawata T, Shimizu Y, Ishizuka T, Dobashi K, Mori M. 4-Hydroxy-2-nonenal IMR-90 மனித நுரையீரல் ஃபைப்ரோபிளாஸ்ட்களால் ஃபைப்ரோனெக்டின் உற்பத்தியை மேம்படுத்துகிறது. ஒழுங்குபடுத்தப்பட்ட கைனேஸ் p44/42 பாதை. டாக்ஸிகோல் ஆப்பிள் பார்மகோல். 2002;184:127-135.
117. Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Kharitonov SA, Barnes PJ. சிஓபிடி மற்றும் ஆரோக்கியமான புகைப்பிடிப்பவர்களுக்கு நுரையீரல் ஆக்ஸிஜனேற்ற அழுத்தத்தின் இன் விவோ பயோமார்க்கராக 8-ஐசோப்ரோஸ்டேன் வெளியேற்றப்படுகிறது. ஆம் ஜே ரெஸ்பிர் கிரிட் கேர் மெட். 2000;162:1175-1177.
118. Morrison D, Rahman I, Lannan S, MacNee W. புகைப்பிடிப்பவர்களின் காற்று இடைவெளிகளில் எபிடெலியல் ஊடுருவல், வீக்கம் மற்றும் ஆக்ஸிஜனேற்ற அழுத்தம். ஆம் ஜே ரெஸ்பிர் கிரிட் கேர் மெட். 1999;159:473-479.
119. Nowak D, Kasielski M, Antczak A, Pietras T, Bialasiewicz P. நிலையான நாள்பட்ட தடுப்பு நுரையீரல் நோயால் பாதிக்கப்பட்ட நோயாளிகளின் காலாவதியான மூச்சுத் திணறலில் தியோபார்பிட்யூரிக் அமிலம்-எதிர்வினை பொருட்கள் மற்றும் ஹைட்ரஜன் பெராக்சைட்டின் அதிகரித்த உள்ளடக்கம்: சிகரெட் புகைப்பதால் குறிப்பிடத்தக்க விளைவு இல்லை. சுவாச மருத்துவம். 1999;93:389-396.
120. கெல்லி FJ, Mudway IS. காற்று-நுரையீரல் இடைமுகத்தில் புரத ஆக்சிஜனேற்றம். அமினோ அமிலங்கள். 2003;25:375-396.
121. டீன் ஆர்டி, ராபர்ட்ஸ் சிஆர், ஜெஸ்ஸப் டபிள்யூ Prog Clin Biol Res. 1985;180:341-350.
122. கெக் ஆர்ஜி. புரதங்களில் மெத்தியோனைன் ஆக்சிஜனேற்றத்திற்கான ஆய்வாக டி-பியூட்டில் ஹைட்ரோபெராக்சைடைப் பயன்படுத்துதல். குத உயிர்வேதியியல். 1996;236:56-62.
123. டேவிஸ் கே.ஜே. ஆக்ஸிஜன் ரேடிக்கல்களால் புரத சேதம் மற்றும் சிதைவு. I. பொது அம்சங்கள். ஜே பயோல் கெம். 1987;262:9895-9901.
124. ஸ்டாட்மேன் ஈஆர். புரதங்களின் உலோக அயனி-வினையூக்கி ஆக்சிஜனேற்றம்: உயிர்வேதியியல் பொறிமுறை மற்றும் உயிரியல் விளைவுகள். இலவச ரேடிக் பயோல் மெட்.
1990;9:315–325.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. கலப்பு-செயல்பாட்டு ஆக்சிஜனேற்ற எதிர்வினைகள் மூலம் முக்கிய வளர்சிதை மாற்ற நொதிகளை செயலிழக்கச் செய்தல்: புரதம் விற்றுமுதல் மற்றும் முதுமையில் சாத்தியமான தாக்கம். Proc Natl Acad Sci US A. 1983;80:1521–1525.
126. Stadtman ER, Moskovitz J, Levine RL. புரதங்களின் மெத்தியோனைன் எச்சங்களின் ஆக்சிஜனேற்றம்: உயிரியல் விளைவுகள். ஆன்டிஆக்சிட் ரெடாக்ஸ் சிக்னல். 2003;5:577-582.
127. ஸ்டாட்மேன் ஈஆர், லெவின் ஆர்எல். இலவச அமினோ அமிலங்கள் மற்றும் புரதங்களில் உள்ள அமினோ அமில எச்சங்களின் ஃப்ரீ ரேடிக்கல்-மத்தியஸ்த ஆக்சிஜனேற்றம். அமினோ அமிலங்கள். 2003;25:207-218.
128. ஸ்டாட்மேன் ஈஆர். வயதான மற்றும் வயது தொடர்பான நோய்களில் புரத ஆக்சிஜனேற்றம். Ann NY Acad Sci. 2001;928:22-38.
129. ஷேக்டர் ஈ. உயிரியல் மாதிரிகளில் புரத ஆக்சிஜனேற்றத்தின் அளவு மற்றும் முக்கியத்துவம். மருந்து மெட்டாப் ரெவ். 2000;32:307–326.
130. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. ஆக்ஸிஜனேற்ற அழுத்தம் மற்றும் செல் சமிக்ஞை. கர்ர் மெட் செம். 2004;11:1163-1182.
131. Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. வாஸ்குலர் எண்டோடெலியல் வளர்ச்சி காரணி (VEGF) மற்றும் அதன் ஏற்பிகள். FASEB J. 1999;13:9-22.
132. சுந்தரேசன் எம், யூ இசட்எக்ஸ், ஃபெரான்ஸ் விஜே, சுல்சினர் டிஜே, குட்கிண்ட் ஜேஎஸ், மற்றும் பலர். ரேக்1 மூலம் ஃபைப்ரோபிளாஸ்ட்களில் எதிர்வினை-ஆக்ஸிஜன் இனங்கள் உருவாக்கத்தை ஒழுங்குபடுத்துதல். பயோகெம் ஜே. 1996;318:379-382.
133. சன் டி, ஓபர்லி எல்டபிள்யூ. டிரான்ஸ்கிரிப்ஷனல் ஆக்டிவேட்டர்களின் ரெடாக்ஸ் ஒழுங்குமுறை. இலவச ரேடிக் பயோல் மெட். 1996;21:335-348.
134. கிளாட் பி, மோலினா இபி, டி லகோபா எம்ஜி, பாடிலா சிஏ, மார்டினெஸ்-கலேஸ்டியோ இ, பார்செனா ஜேஏ, லாமாஸ் எஸ். ரிவர்சிபிள் எஸ்-குளுதாதியோலேஷன் மூலம் சி-ஜூன் டிஎன்ஏ பிணைப்பின் ரெடாக்ஸ் ஒழுங்குமுறை. FASEB J. 1999;13:1481-1490.
135. Reynaert NL, Ckless K, Guala AS, Wouters EF, van der Vliet A, Janssen Heininger
YM குளுடரெடாக்சின்-1 வினையூக்கிய சிஸ்டைன் வழித்தோன்றலைத் தொடர்ந்து எஸ்-குளுதாதியோனிலேட்டட் புரதங்களை சிட்டு கண்டறிதல். பயோகிம் பயோஃபிஸ் ஆக்டா. 2006;1760:380-387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. குளுடரெடாக்சின்-1 மாடுலேஷன்
ஒவ்வாமை காற்றுப்பாதை நோயின் சுட்டி மாதிரியில் வெளிப்பாடு. ஆம் ஜே ரெஸ்பிர் செல் மோல் பயோல். 2007;36:147-151.
137. ஃபிலோமெனி ஜி, ரோட்டிலியோ ஜி, சிரியோலோ எம்ஆர். செல் சிக்னலிங் மற்றும் குளுதாதயோன் ரெடாக்ஸ் அமைப்பு. பயோகெம் பார்மகோல். 2002;64:1057-1064.
138. பாண்டே வி, ராமோஸ் எம்.ஜே. அணுக்கரு காரணி-கப்பா பி மற்றும் பிற செல்லுலார் புரதங்களால் 15-டியோக்ஸைடெல்டா (12,14) புரோஸ்டாக்லாண்டின் ஜே(2) மூலக்கூறு அங்கீகாரம். Bioorg மெட் செம் லெட். 2005;15:4057–4063.
139. பெர்கின்ஸ் என்டி. NF-kappaB மற்றும் IKK செயல்பாட்டுடன் செல்-சிக்னலிங் பாதைகளை ஒருங்கிணைத்தல். நாட் ரெவ் மோல் செல் பயோல். 2007;8:49-62.
140. கில்மோர் டிடி. NF-kappaB அறிமுகம்: வீரர்கள், பாதைகள், முன்னோக்குகள். புற்றுநோயியல். 2006;25:6680–6684.
141. Hirota K, Murata M, Sachi Y, Nakamura H, Takeuchi J, Mori K, Yodoi J. சைட்டோபிளாசம் மற்றும் நியூக்ளியஸில் தியோரெடாக்ஸின் தனித்துவமான பாத்திரங்கள். டிரான்ஸ்கிரிப்ஷன் காரணி NF-kappaB இன் ரெடாக்ஸ் ஒழுங்குமுறையின் இரண்டு-படி வழிமுறை. ஜே பயோல் கெம். 1999;274:27891-27897.
142. வார்டு PA. கடுமையான நுரையீரல் காயத்தில் நிரப்பு, கெமோக்கின்கள் மற்றும் ஒழுங்குமுறை சைட்டோகைன்களின் பங்கு. Ann NY Acad Sci. 1996;796:104-112.
143. சைட்டோகைன் மரபணு ஒழுங்குமுறையில் அகிரா எஸ், கிஷிமோட்டோ ஏ. என்எஃப்-ஐஎல்6 மற்றும் என்எஃப்-கேபி. Adv Immunol. 1997;65:1-46.
144. மேயர் எம், ஷ்ரெக் ஆர், பேயுர்லே பிஏ. H2O2 மற்றும் ஆன்டிஆக்ஸிடன்ட்கள் NF-kappa B மற்றும் AP-1 ஆகியவற்றை அப்படியே செல்களில் செயல்படுத்துவதில் எதிர் விளைவுகளைக் கொண்டுள்ளன: AP-1 இரண்டாம் நிலை ஆக்ஸிஜனேற்ற-பதிலளிக்கக்கூடிய காரணியாக உள்ளது. EMBO J. 1993;12:2005-2015.
145. அபேட் சி, படேல் எல், ரௌஷர் எஃப்ஜே, கர்ரான் டி. ஃபோஸின் ரெடாக்ஸ் ஒழுங்குமுறை மற்றும் விட்ரோவில் ஜூன் டிஎன்ஏ-பைண்டிங் செயல்பாடு. விஞ்ஞானம். 1990;249:1157-1161.
146. Galter D, Mihm S, Droge W. அணுக்கரு டிரான்ஸ்கிரிப்ஷன் காரணிகள் kB மற்றும் ஆக்டிவேட்டர் புரோட்டீன்-1 மீது குளுதாதயோன் டிசல்பைட்டின் தனித்துவமான விளைவுகள். யூர் ஜே பயோகெம். 1994;221:639-648.
147. Hirota K, Matsui M, Iwata S, Nishiyama A, Mori K, Yodoi J. AP-1 டிரான்ஸ்கிரிப்ஷனல் செயல்பாடு தியோரெடாக்சின் மற்றும் Ref-1 ஆகியவற்றுக்கு இடையே உள்ள நேரடித் தொடர்பு மூலம் கட்டுப்படுத்தப்படுகிறது. Proc Natl Acad Sci US A. 1997;94: 3633–3638.
IFM இன் ஃபைண்ட் எ பிராக்டிஷனர் கருவி என்பது செயல்பாட்டு மருத்துவத்தில் மிகப்பெரிய பரிந்துரை வலையமைப்பாகும், இது நோயாளிகளுக்கு உலகில் எங்கிருந்தும் செயல்பாட்டு மருத்துவப் பயிற்சியாளர்களைக் கண்டறிய உதவுவதற்காக உருவாக்கப்பட்டது. IFM சான்றளிக்கப்பட்ட பயிற்சியாளர்கள் தேடல் முடிவுகளில் முதலில் பட்டியலிடப்பட்டுள்ளனர், அவர்கள் செயல்பாட்டு மருத்துவத்தில் விரிவான கல்வியைப் பெற்றுள்ளனர்